Eukaryoter har to hovedtyper af celledeling: mitose og meiose. Mitose bruges til at producere nye kropsceller til vækst og heling, mens meiose bruges til at producere kønsceller (æg og sædceller). Meiose vil blive behandlet i et senere kapitel.
Cellecyklus er en ordnet række af begivenheder, der involverer cellevækst og celledeling, som producerer to nye datterceller via mitose. Længden af cellecyklusen er meget variabel, selv inden for cellerne i en enkelt organisme. Hos mennesker varierer hyppigheden af celleomsætningen fra et par timer i den tidlige embryonale udvikling til et gennemsnit på to til fem dage for epitelceller eller til et helt menneskeliv, der tilbringes uden at dele sig i specialiserede celler som f.eks. kortikale neuroner eller hjertemuskelceller. Der er også variation i den tid, som en celle tilbringer i hver fase af cellecyklussen. Når hurtigt delende pattedyrsceller dyrkes i kultur (uden for kroppen under optimale vækstbetingelser), er cyklusens længde ca. 24 timer. Tidspunktet for begivenhederne i cellecyklussen styres af mekanismer, der er både interne og eksterne i forhold til cellen.
Celler på vej mod celledeling går gennem en række præcist timede og nøje regulerede faser af vækst, DNA-replikation og deling, der producerer to genetisk identiske celler. Cellecyklussen har to hovedfaser: interfasen og den mitotiske fase (figur 1). I interfasen vokser cellen, og DNA’et replikeres. I den mitotiske fase adskilles det replikerede DNA og cytoplasmaindholdet, og cellen deler sig.
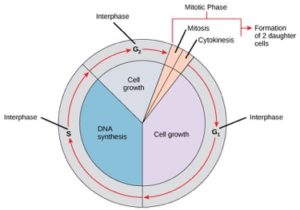
I interfasen gennemgår cellen normale processer, samtidig med at den forbereder sig på celledeling. For at en celle kan gå fra interfasen til mitosefasen, skal mange interne og eksterne betingelser være opfyldt. De tre stadier i interfasen kaldes G1, S og G2.
G1-fasen (første hul)
Den første fase i interfasen kaldes G1-fasen (første hul), fordi der fra et mikroskopisk synspunkt kun er få ændringer synlige. I G1-stadiet er cellen imidlertid ret aktiv på det biokemiske plan. Cellen ophober byggestenene til kromosom-DNA og de tilhørende proteiner samt ophober tilstrækkelige energireserver til at fuldføre opgaven med at replikere hvert kromosom i kernen.
S-fase (DNA-syntese)
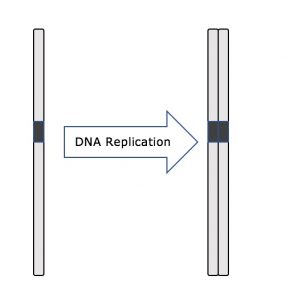
I hele interfasen forbliver kerne-DNA’et i en halvkondenseret kromatinkonfiguration. I S-fasen kan DNA-replikationen fortsætte gennem de mekanismer, der resulterer i dannelsen af identiske par af DNA-molekyler – søsterkromatider – som er fast knyttet til det centromeriske område (figur 2).
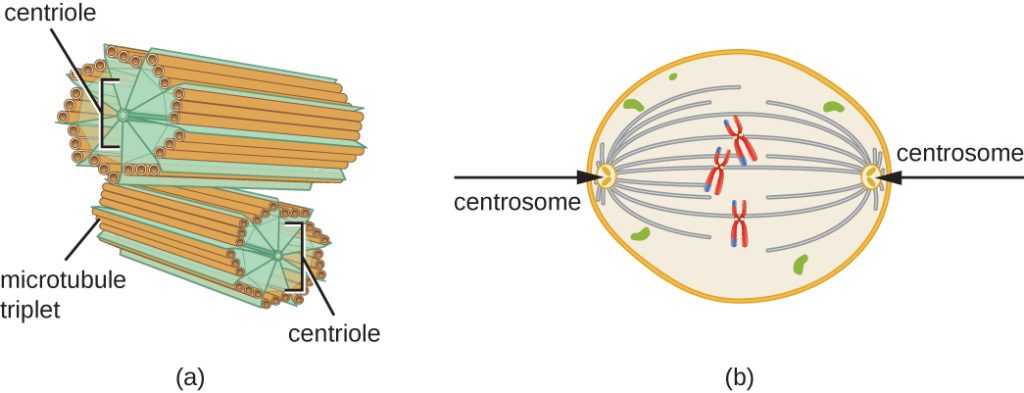
Centrosomet duplikeres også i løbet af S-fasen. De to centrosomer vil give anledning til den mitotiske spindel, det apparat, der orkestrerer kromosomernes bevægelse under mitosen. I centrum af hver dyrecelle er centrosomerne i dyreceller forbundet med et par stavlignende objekter, centriolerne, som står vinkelret på hinanden. Centrioler er med til at organisere celledelingen. Centrioler er ikke til stede i centrosomerne hos andre eukaryote arter, såsom planter og de fleste svampe.
G2-fasen (andet hul)
I G2-fasen genopfylder cellen sine energidepoter og syntetiserer proteiner, der er nødvendige for kromosommanipulation. Nogle celleorganeller duplikeres, og cytoskelettet nedbrydes for at skaffe ressourcer til mitosefasen. Der kan forekomme yderligere cellevækst i G2-perioden. De sidste forberedelser til den mitotiske fase skal være afsluttet, før cellen kan gå ind i mitosens første fase.
Den mitotiske fase
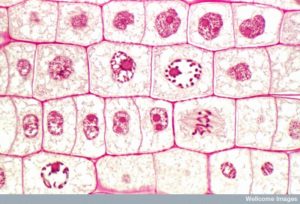
For at lave to datterceller skal indholdet af kernen og cytoplasmaet deles. Mitosefasen er en proces i flere trin, hvor de duplikerede kromosomer bliver justeret, adskilt og flyttet til modsatte poler i cellen, hvorefter cellen deles i to nye identiske datterceller. Den første del af den mitotiske fase, mitose, består af fem faser, som gennemfører kerneopdelingen (figur 5). Den anden del af mitosefasen, kaldet cytokinese, er den fysiske adskillelse af cytoplasmabestanddelene i to datterceller. Selv om mitosefaserne ligner hinanden for de fleste eukaryoter, er cytokineseprocessen helt anderledes for eukaryoter, der har cellevægge, som f.eks. planteceller.
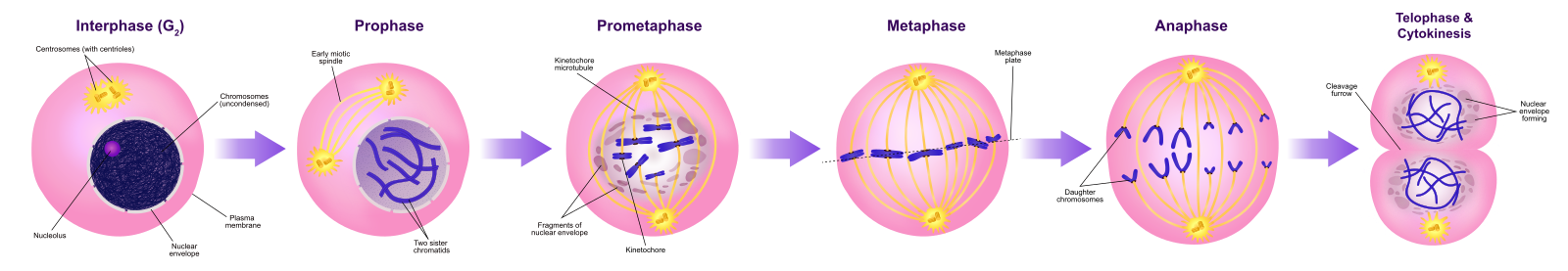
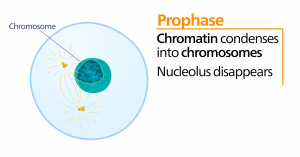
Profasen
Under profasen, den “første fase”, begynder kernehyllen at dissocieres i små vesikler, og de membranøse organeller (såsom Golgi-apparatet og det endoplasmatiske retikulum), fragmenteres og spredes mod cellens kanter. Nukleolus forsvinder. Centrosomerne begynder at bevæge sig til cellens modsatte poler. Mikrotubuli, som vil danne mitotisk spindel, strækker sig mellem centrosomerne og skubber dem længere fra hinanden, efterhånden som mikrotubulifibrene bliver længere. Søsterkromatiderne begynder at vikle sig mere stramt sammen ved hjælp af kondensinproteiner og bliver synlige i et lysmikroskop.
Prometafase
Under prometafase, den “første forandringsfase”, fortsætter mange processer, der blev påbegyndt i profasen, med at skride fremad. Resterne af kernehyllen fragmenterer. Den mitotiske spindel fortsætter med at udvikle sig, efterhånden som flere mikrotubuli samles og strækker sig over længden af det tidligere kerneområde. Kromosomerne bliver mere kondenserede og diskrete. Hver søsterkromatid udvikler en proteinstruktur kaldet en kinetokore i det centromeriske område.
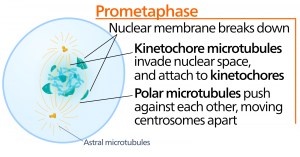
Proteinerne i kinetochoren tiltrækker og binder mitotiske spindelmikrotubuli til mitotiske spindelmikrotubuli. Når spindelmikrotubuli strækker sig fra centrosomerne, kommer nogle af disse mikrotubuli i kontakt med og binder sig fast til kinetokorerne. Når først en mitotisk fiber hæfter sig på et kromosom, vil kromosomet blive orienteret, indtil søsterkromatidernes kinetochorer vender mod de modsatte poler. Til sidst vil alle søsterkromatider være knyttet via deres kinetokorer til mikrotubuli fra modsatrettede poler. Spindelmikrotubuli, der ikke griber ind i kromosomerne, kaldes polære mikrotubuli. Disse mikrotubuli overlapper hinanden midtvejs mellem de to poler og bidrager til cellens strækning. Astralmikrotubuli er placeret nær polerne, hjælper med spindelorienteringen og er nødvendige for reguleringen af mitose.

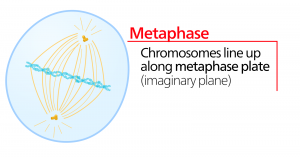
Metafase
Under metafase, “forandringsfasen”, er alle kromosomer rettet ud i et plan kaldet metafasepladen eller det ækvatoriale plan, midt mellem cellens to poler. Søsterkromatiderne er stadig tæt knyttet til hinanden af cohesinproteiner. På dette tidspunkt er kromosomerne maksimalt kondenseret.
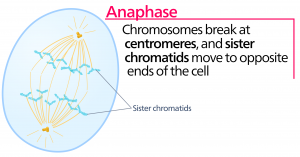
Anafasen
Under anafasen, den “opadgående fase”, nedbrydes cohesinproteinerne, og søsterkromatiderne adskilles ved centromeren. Hver kromatid, som nu kaldes et kromosom, trækkes hurtigt mod centrosomet, som dets mikrotubulus er fastgjort til. Cellen bliver synligt forlænget (ovalformet), da de polære mikrotubuli glider mod hinanden ved metafasepladen, hvor de overlapper hinanden.
Telofase
Under telofase, “afstandsfasen”, når kromosomerne de modsatte poler og begynder at dekondensere (udtrævle) og slapper af i en kromatin-konfiguration. De mitotiske spindler depolymeriseres til tubulin-monomerer, som vil blive brugt til at samle cytoskeletale komponenter til hver dattercelle. Der dannes kerneomslag omkring kromosomerne, og nukleosomer fremkommer inden for kerneområdet.
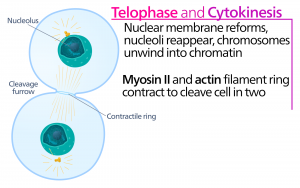
Cytokinese
Cytokinese, eller “cellebevægelse”, er det andet hovedstadie i mitosefasen, hvor celledelingen afsluttes via den fysiske adskillelse af cytoplasmabestanddelene i to datterceller. Delingen er ikke afsluttet, før cellekomponenterne er blevet delt og fuldstændig adskilt i de to datterceller. Selv om mitosefaserne ligner hinanden for de fleste eukaryoter, er processen med cytokinese helt anderledes for eukaryoter, der har cellevægge, som f.eks. planteceller.
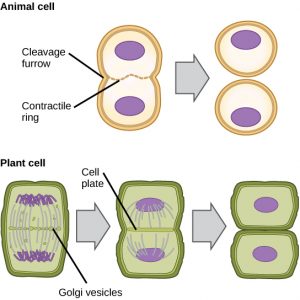
I celler som f.eks. dyreceller, der mangler cellevægge, følger cytokinese efter starten af anafase. Der dannes en kontraktile ring bestående af aktinfilamenter lige inden for plasmamembranen ved den tidligere metafaseplade (Figur 12). Aktinfilamenterne trækker cellens ækvator indad og danner en spalte. Denne revne, eller “spalte”, kaldes spaltningsfuren. Furen bliver dybere, efterhånden som aktinringen trækker sig sammen, og til sidst spaltes membranen i to.
I planteceller skal der dannes en ny cellevæg mellem dattercellerne. Under interfasen ophober Golgi-apparatet enzymer, strukturelle proteiner og glukosemolekyler, inden det deles i vesikler og spredes ud over hele den delende celle (figur 12). I telofase transporteres disse Golgi-vesikler på mikrotubuli for at danne en phragmoplast (en vesikulær struktur) på metafasepladen. Her smelter vesiklerne sammen og samler sig fra midten mod cellevæggene; denne struktur kaldes en celleplade. Efterhånden som flere vesikler smelter sammen, udvider cellepladen sig, indtil den smelter sammen med cellevæggene i cellens periferi. Enzymer bruger den glukose, der har ophobet sig mellem membranlagene, til at opbygge en ny cellevæg. Golgimembranerne bliver dele af plasmamembranen på hver side af den nye cellevæg.
Summary of Mitosis and Cytokinesis
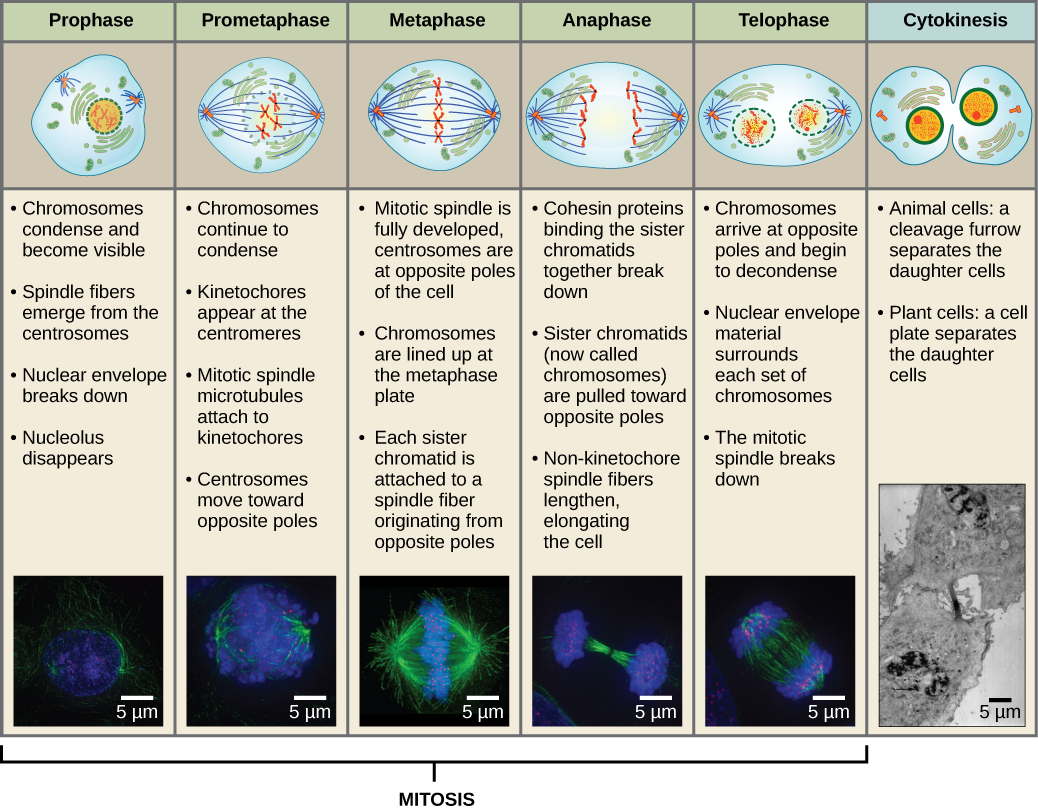
Figur 13 Mitose er opdelt i fem stadier – profasen, prometafasen, metafasen, anafasen og telofasen. Billederne nederst er taget ved fluorescensmikroskopi af celler, der er kunstigt farvet med fluorescerende farvestoffer: blå fluorescens angiver DNA (kromosomer) og grøn fluorescens angiver mikrotubuli (spindelapparat). (kredit “mitosis drawings”: ændring af Mariana Ruiz Villareal; kredit “micrographs”: ændring af Roy van Heesbeen; kredit “cytokinesis micrographs”: Wadsworth Center/New York State Department of Health; skala-bar data fra Matt Russell)
G0-fase
Det er ikke alle celler, der følger det klassiske cellecyklusmønster, hvor en nyoprettet dattercelle straks går ind i interfasen, tæt fulgt af mitosefasen. Celler i G0-fasen forbereder sig ikke aktivt på at dele sig. Cellen befinder sig i en hvilende (inaktiv) fase, idet den har forladt cellecyklusen. Nogle celler går midlertidigt ind i G0-fasen, indtil et eksternt signal udløser indtræden af G1-fasen. Andre celler, der aldrig eller sjældent deler sig, f.eks. modne hjertemuskel- og nerveceller, forbliver permanent i G0).