Evolution og naturlig udvælgelse
Naturen tillader ingen løssluppenhed, undskylder ingen fejl
– Ralph Waldo Emerson
Jeg har kaldt dette princip, hvorefter enhver lille variation, hvis den er nyttig, bevares, for naturlig udvælgelse.
– Charles Darwin, Arternes oprindelse
|
10 okt 2010 |
|
I denne lektion ønsker vi at spørge:
- Hvordan førte iagttagelser i naturen til formuleringen af evolutionsteorien?
- Hvad er hovedpunkterne i Darwins evolutionsteori?
- Hvordan fungerer den naturlige udvælgelsesproces?
- Hvilke beviser har vi for lokal tilpasning?
- Hvordan kan naturlig udvælgelse påvirke frekvensen af egenskaber i løbet af på hinanden følgende generationer?
Teoriens (R)evolution
Evolutionsteorien er en af de store intellektuelle revolutioner i menneskehedens historie, som drastisk har ændret vores opfattelse af verden og vores plads i den. Charles Darwin fremlagde en sammenhængende evolutionsteori og samlede en lang række beviser til støtte for denne teori. På Darwins tid troede de fleste videnskabsmænd fuldt ud på, at hver organisme og hver tilpasning var skabt af skaberen. Linneaus etablerede det biologiske klassifikationssystem, som vi bruger i dag, og han gjorde det i en ånd af katalogisering af Guds skabelser.
Med andre ord blev alle ligheder og uligheder mellem grupper af organismer, som er resultatet af den forgreningsproces, der skaber livets store træ (se figur 1), af filosoffer og videnskabsmænd fra det tidlige 19. århundrede betragtet som en konsekvens af et almægtigt design.
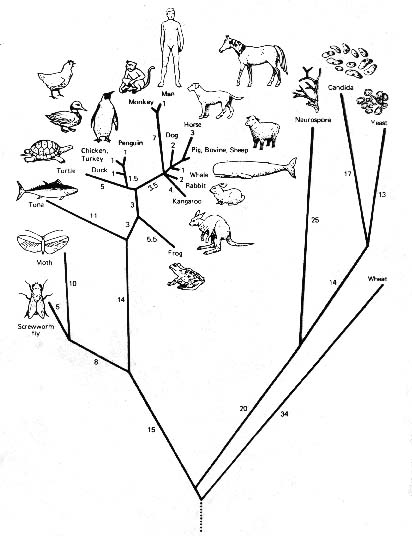
Figur 1: Et fylogenetisk “livstræ” konstrueret ved computeranalyse af cyochrom c-molekyler i de viste organismer; der findes lige så mange forskellige livstræer, som der findes analysemetoder til at konstruere dem.
I det 19. århundrede var en række naturhistorikere imidlertid begyndt at tænke på evolutionære forandringer som en forklaring på de mønstre, der observeres i naturen. Følgende ideer var en del af det intellektuelle klima på Darwins tid.
- Ingen vidste, hvor gammel jorden var, men geologer var begyndt at lave skøn over, at jorden var betydeligt ældre end forklaret af den bibelske skabelse. Geologerne lærte mere om lag, eller lag, der er dannet af successive perioder med aflejring af sedimenter. Dette tydede på en tidsmæssig rækkefølge, hvor yngre lag overlejrede ældre lag.
- Et koncept kaldet uniformitarisme, som i høj grad skyldes den indflydelsesrige geolog Charles Lyell, gik ud på at tyde jordens historie ud fra den arbejdshypotese, at de nuværende forhold og processer er nøglen til fortiden, ved at undersøge igangværende, observerbare processer som erosion og aflejring af sedimenter.
- Opdagelserne af fossiler hobede sig op i løbet af det 18. og 19. århundrede. I begyndelsen troede naturforskere, at de fandt rester af ukendte, men stadig levende arter. Efterhånden som fossilfundene fortsatte, blev det imidlertid klart, at man ikke kendte noget, der lignede kæmpedinosaurer, fra noget sted på planeten. Desuden påpegede Cuvier allerede i 1800, at jo dybere lagene var, jo mindre lignede fossilerne eksisterende arter.
- Ligheder mellem grupper af organismer blev anset for at være tegn på slægtskab, hvilket igen tydede på evolutionære forandringer. Darwins intellektuelle forgængere accepterede ideen om evolutionære relationer mellem organismer, men de kunne ikke give en tilfredsstillende forklaring på, hvordan evolutionen fandt sted.
- Lamarck er den mest berømte af disse. I 1801 foreslog han organisk evolution som forklaring på den fysiske lighed mellem grupper af organismer, og han foreslog en mekanisme for adaptiv forandring baseret på nedarvning af erhvervede egenskaber. Han skrev om giraffen:
“Vi ved, at dette dyr, det højeste af pattedyrene, lever i det indre af Afrika, på steder, hvor jorden, der næsten altid er tør og uden græs, tvinger det til at æde træer og til hele tiden at anstrenge sig for at nå dem. Denne vane, der er blevet opretholdt i lang tid, har hos alle medlemmer af dens race haft det resultat, at forbenene er blevet længere end bagbenene, og at dens hals er blevet så strakt, at giraffen, uden at stå på bagbenene, løfter hovedet til en højde af seks meter.”
Dette siger i bund og grund, at giraffernes hals er blevet lang som følge af, at de hele tiden strækker sig for at nå højt løvværk. Larmarck var naturligvis ukorrekt i den antagne mekanisme, men hans eksempel gør det klart, at naturforskere tænkte over muligheden for evolutionære forandringer i begyndelsen af 1800-tallet.
- Darwin blev påvirket af observationer, der blev gjort under hans ungdomsrejse som naturforsker på undersøgelsesskibet Beagle. På Galapagosøerne bemærkede han de små variationer, der gjorde skildpadder fra forskellige øer genkendeligt forskellige. Han observerede også en hel række unikke finker, de berømte “Darwins finker”, som udviste små forskelle fra ø til ø. Desuden syntes de alle at ligne, men adskille sig fra den almindelige finke på Ecuadors fastland, 600 miles mod øst. Mønstre i organismers udbredelse og lighed havde en vigtig indflydelse på Darwins tankegang. Billedet øverst på denne side er af Darwins egne skitser af finker i hans Journal of Researches.
- I 1859 udgav Darwin sin berømte bog On the Origin of Species by Means of Natural Selection, et bind på over 500 sider, der samlede omfattende beviser for sin teori. Udgivelsen af bogen vakte furore – alle eksemplarer af bogen blev solgt samme dag, som den udkom. Medlemmer af det religiøse samfund samt nogle videnskabelige kolleger var forargede over Darwins idéer og protesterede. De fleste videnskabsmænd anerkendte imidlertid styrken i Darwins argumenter. I dag diskuterer skolebestyrelser stadig gyldigheden og egnetheden af Darwins teori i de naturvidenskabelige læseplaner, og der er opstået en hel debat omkring kontroversen (se WWW-siden Talk.Origins for en løbende dialog). Vi har ikke tid til at gennemgå alle Darwins beviser og argumenter, men vi kan undersøge de centrale idéer. Hvad siger denne evolutionsteori?
Darwins teori
Darwins evolutionsteori indebærer følgende grundlæggende ideer. De tre første ideer var allerede til diskussion blandt tidligere og samtidige naturforskere, der arbejdede med “artsproblemet”, da Darwin begyndte sin forskning. Darwins oprindelige bidrag var mekanismen for naturlig udvælgelse og rigelige mængder af beviser for evolutionære ændringer fra mange kilder. Han gav også gennemtænkte forklaringer på konsekvenserne af evolutionen for vores forståelse af livets historie og den moderne biologiske mangfoldighed.
-
Species (populationer af krydsende organismer) ændrer sig over tid og rum. Repræsentanterne for de arter, der lever i dag, adskiller sig fra dem, der levede i den seneste tid, og populationer i forskellige geografiske områder i dag adskiller sig lidt i form eller adfærd. Disse forskelle strækker sig ind i de fossile optegnelser, som giver rigelig støtte til denne påstand.
-
Alle organismer deler fælles forfædre med andre organismer. Med tiden kan populationer dele sig op i forskellige arter, som deler en fælles forfædres population. Hvis man går langt nok tilbage i tiden, har ethvert par organismer en fælles forfader. For eksempel delte mennesker en fælles forfader med chimpanser for ca. otte millioner år siden, med hvaler for ca. 60 millioner år siden og med kænguruer for over 100 millioner år siden. Fælles forfædre forklarer lighederne mellem organismer, der er klassificeret sammen: Deres ligheder afspejler arven af træk fra en fælles forfader.
-
Evolutionære forandringer er gradvise og langsomme efter Darwins opfattelse. Denne påstand blev understøttet af de lange episoder af gradvise forandringer hos organismer i de fossile optegnelser og af det faktum, at ingen naturforsker havde observeret den pludselige fremkomst af en ny art på Darwins tid. Siden da har biologer og palæontologer dokumenteret et bredt spektrum af langsomme til hurtige hastigheder af evolutionære ændringer inden for slægter.
Den primære mekanisme for forandring over tid er naturlig selektion, som uddybes nedenfor. Denne mekanisme forårsager ændringer i organismers egenskaber (karaktertræk) inden for slægter fra generation til generation.
Darwins proces for naturlig selektion
Darwins proces for naturlig selektion har fire komponenter.
- Variation. Organismer (inden for populationer) udviser individuel variation i udseende og adfærd. Disse variationer kan omfatte kropsstørrelse, hårfarve, ansigtsmærker, stemmeegenskaber eller antallet af afkom. På den anden side viser nogle egenskaber kun lidt eller ingen variation mellem individer – f.eks. antallet af øjne hos hvirveldyr.
- Arv. Nogle egenskaber går konsekvent i arv fra forældrene til afkommet. Sådanne egenskaber er arvelige, mens andre egenskaber er stærkt påvirket af miljøforhold og har en svag arvelighed.
- Høj vækstrate i populationen. De fleste populationer får flere afkom hvert år, end de lokale ressourcer kan bære, hvilket fører til en kamp om ressourcerne. Hver generation oplever en betydelig dødelighed.
- Differentieret overlevelse og reproduktion. Individer, der har egenskaber, der er velegnede til kampen om de lokale ressourcer, vil bidrage med flere afkom til den næste generation.
Fra den ene generation til den næste vil kampen om ressourcerne (det Darwin kaldte “kampen for eksistens”) favorisere individer med nogle variationer frem for andre og derved ændre frekvensen af træk i populationen. Denne proces er naturlig selektion. De træk, der giver en fordel til de individer, der efterlader flere afkom, kaldes tilpasninger.
For at naturlig selektion kan virke på et træk, skal det pågældende træk besidde arvelig variation og give en fordel i konkurrencen om ressourcerne. Hvis et af disse krav ikke er til stede, oplever egenskaben ikke naturlig selektion. (Vi ved nu, at sådanne træk kan ændre sig ved andre evolutionære mekanismer, som er blevet opdaget siden Darwins tid).
Naturlig selektion opererer efter komparative fordele, ikke efter en absolut standard for design. “…da naturlig udvælgelse virker ved konkurrence om ressourcer, tilpasser den kun indbyggerne i hvert land i forhold til graden af perfektion hos deres medmennesker” (Charles Darwin, On the Origin of Species, 1859).
I løbet af det 20. århundrede blev genetik integreret med Darwins mekanisme, hvilket gør det muligt for os at vurdere naturlig udvælgelse som den differentierede overlevelse og reproduktion af genotyper, der svarer til bestemte fænotyper. Naturlig selektion kan kun virke på eksisterende variation inden for en population. Sådanne variationer opstår ved mutation, en ændring i en del af den genetiske kode for en egenskab. Mutationer opstår tilfældigt og uden forudseenhed for den potentielle fordel eller ulempe ved mutationen. Med andre ord opstår variationer ikke, fordi der er behov for dem.
Bevis for naturlig selektion
Lad os se på et eksempel for at gøre den naturlige selektion tydelig.
Industriel melanisme er et fænomen, der ramte over 70 arter af møl i England. Det er blevet bedst undersøgt hos pebermøl, Biston betularia. Før 1800 havde det typiske møl af denne art et lyst mønster (se figur 2). Mørkfarvede eller melaniske møl var sjældne og var derfor samlerobjekter.

Figur 2. Billede af pebermøl
Under den industrielle revolution formørkede sod og andet industrielt affald træstammer og dræbte laver. Den lyse morph af møl blev sjælden, og den mørke morph blev rigeligt udbredt. I 1819 blev den første melaniske morph set; i 1886 var den langt mere almindelig – hvilket illustrerer en hurtig evolutionær forandring. 
Entlig var lyse morfer kun almindelige på nogle få lokaliteter, langt fra industriområder. Årsagen til denne forandring blev anset for at være selektiv prædation fra fugle, som favoriserede camouflagefarve hos mølet.
I 1950’erne foretog biologen Kettlewell forsøg med udsætning og genfangst med begge morfer. Et kort resumé af hans resultater er vist nedenfor. Ved at observere fuglenes prædation fra persienner kunne han bekræfte, at mølets iøjnefaldende karakter havde stor indflydelse på chancen for, at det ville blive ædt.
Succes med genfangst
|
lys møl |
mørk møl |
|
|
ikke-industrielle skove |
14.6 % |
4.7 % |
|
industrielt træ |
13 % |
27.5 % |
Lokal tilpasning – flere eksempler
Så langt har vi i dagens foredrag understreget, at den naturlige selektion er hjørnestenen i evolutionsteorien. Det giver mekanismen for adaptive ændringer. Enhver ændring i miljøet (f.eks. en ændring i baggrundsfarven på den træstamme, som du sidder på) vil sandsynligvis føre til lokal tilpasning. Enhver udbredt population vil sandsynligvis opleve forskellige miljøforhold i forskellige dele af sit udbredelsesområde. Som følge heraf vil den snart bestå af en række delpopulationer, som adskiller sig lidt eller endog betydeligt fra hinanden.
Der er følgende eksempler, der illustrerer befolkningers tilpasning til lokale forhold.
- Ratteslangen, Elaphe obsoleta, har genkendeligt forskellige populationer på forskellige lokaliteter i det østlige Nordamerika (se figur 3). Det kan diskuteres, om disse skal kaldes geografiske “racer” eller underarter. Disse populationer udgør alle én art, fordi der kan ske parring mellem tilstødende populationer, hvilket får arterne til at dele en fælles genpulje (se foredraget om artsdannelse).
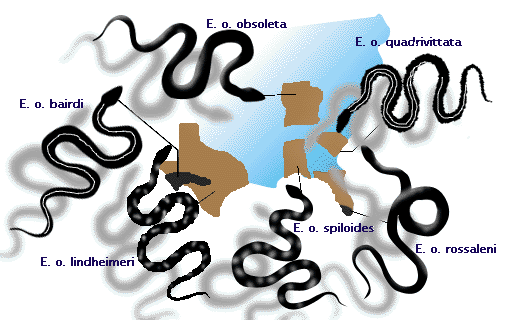
Figur 3: Underarter af rotteslangen Elaphe obsoleta, som krydses med hinanden, hvor deres udbredelsesområder mødes.
- Galapagosfinker er det berømte eksempel fra Darwins rejse. Hver ø på Galapagos, som Darwin besøgte, havde sin egen art af finker (14 i alt), som ikke findes andre steder i verden. Nogle havde næb tilpasset til at æde store frø, andre til små frø, nogle havde papegøje-lignende næb til at æde knopper og frugter, og nogle havde slanke næb til at æde små insekter (se figur 4). En af dem brugte en torn til at undersøge efter insektlarver i træ, ligesom nogle spætter gør. (Seks var jordboere, og otte var træfinker.) (Denne diversificering i forskellige økologiske roller, eller nicher, menes at være nødvendig for at multikarters sameksistens er mulig, et emne, som vi vil undersøge i et senere foredrag). For Darwin viste det sig, at de hver især var let modificerede fra en oprindelig kolonist, sandsynligvis finken på det sydamerikanske fastland, ca. 600 miles mod øst. Det er sandsynligt, at adaptiv stråling førte til dannelsen af så mange arter, fordi andre fugle var få eller fraværende, hvilket efterlod tomme nicher at udfylde, og fordi de mange øer på Galapagos gav rig mulighed for geografisk isolation.
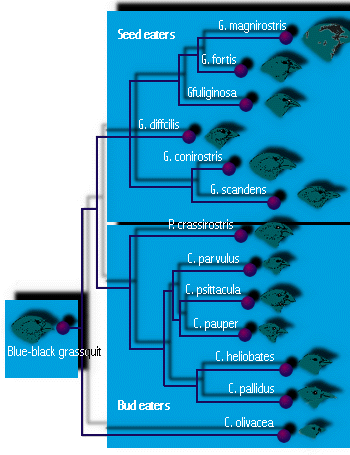
Figur 4
Stabiliserende, retningsbestemt og diversificerende selektion
Sidst vil vi se på en statistisk måde at tænke på selektion på. Lad os antage, at hver population kan beskrives som en frekvensfordeling for en eller anden egenskab — for eksempel næbstørrelse. Bemærk igen, at variation i en egenskab er det afgørende råmateriale for, at evolution kan finde sted.
Hvordan vil frekvensfordelingen se ud i den næste generation?
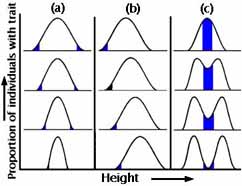
Figurer 5a-c
Først kan andelen af individer med hver værdi af egenskaben (størrelse af næb eller kropsvægt) være nøjagtig den samme. For det andet kan der være retningsbestemt ændring i kun én retning. For det tredje (og med en sådan sjældenhed, at dens eksistens er diskutabel) kan der være samtidige ændringer i begge retninger (f.eks. favoriseres både større og mindre næb på bekostning af dem af mellemstørrelse). Figurerne 5a-c viser disse tre hovedkategorier af naturlig udvælgelse.
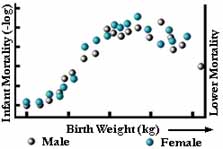
Figur 6
Under stabiliserende selektion elimineres ekstreme varianter fra begge ender af frekvensfordelingen. Frekvensfordelingen ser nøjagtig ud, som den gjorde i generationen før (se figur 5a). Dette er formentlig den mest almindelige form for naturlig selektion, og vi forveksler den ofte med ingen selektion. Et eksempel fra det virkelige liv er fødselsvægten hos menneskebørn (se figur 6).
Under retningsbestemt selektion klarer individer i den ene ende af fordelingen af næbstørrelser sig særligt godt, og derfor forskydes frekvensfordelingen af egenskaben i den efterfølgende generation i forhold til hvor den var i forældregenerationen (se figur 5b). Det er det, vi normalt tænker på som naturlig udvælgelse. Industriel melanisme var et sådant eksempel.

Figur 7
Hestens fossile afstamning giver en bemærkelsesværdig demonstration af retningsbestemt succession. Den fulde slægtslinje er ganske kompliceret og er ikke blot en simpel linje fra den lille morgenhest Hyracotherium fra det tidlige Eocæn til nutidens velkendte Equus. Overordnet set har hesten dog udviklet sig fra en forfader med lille krop, der var bygget til at bevæge sig gennem skove og buske, til dens langbenede efterkommer, der er bygget til at bevæge sig hurtigt på det åbne græsland. Denne udvikling har medført veldokumenterede ændringer i tænder, benlængde og tåstruktur (se figur 7).
Under diversificerende (forstyrrende) selektion favoriseres begge ekstremer på bekostning af de mellemliggende sorter (se figur 5c). Dette er ualmindeligt, men af teoretisk interesse, fordi det antyder en mekanisme for artsdannelse uden geografisk isolation (se foredraget om artsdannelse).
Resumé
Darwins evolutionsteori ændrede fundamentalt retningen for den fremtidige videnskabelige tænkning, selv om den var bygget på en voksende tankegang, der begyndte at sætte spørgsmålstegn ved tidligere ideer om naturens verden.
Kernen i Darwins teori er naturlig selektion, en proces, der sker over på hinanden følgende generationer og defineres som differentiel reproduktion af genotyper.
Naturlig selektion kræver arvelig variation i en given egenskab og differentieret overlevelse og reproduktion i forbindelse med besiddelse af denne egenskab.
Eksempler på naturlig selektion er veldokumenterede, både ved observation og gennem de fossile optegnelser.
Selektion virker på frekvensen af egenskaber, og kan tage form af stabiliserende, retningsbestemt eller diversificerende selektion.
Foreslået læsning
o Darwin, C. 1959. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life (Om arternes opståen ved hjælp af naturlig udvælgelse, eller bevarelse af begunstigede racer i kampen for livet). London: J. Murray.
o Futuyma, D.J. 1986. Evolutionær biologi. Sunderland, Mass: Sinauer Associates, Inc.
o Dawkins, R. 1989. The Selfish Gene. Oxford: Oxford University Press.