Introduktion
Charofytter eller basale streptofytter; (Becker og Marin, 2009; Leliaert et al.., 2012) udgør et mangfoldigt taksonomisk sortiment af eksisterende ferskvands- og terrestriske grønne alger, der viser en bred vifte af encellede, filamentøse og “parenchymatøse” former (Graham, 1993; Lewis og McCourt, 2004). En forfædres slægt af charofytter dukkede op på og koloniserede land for 450-500 millioner år siden. Disse organismer tilpassede sig terrestriske forhold, blev i stand til at overleve og reproducere, når de var fuldt eksponeret for atmosfæren, og nogle medlemmer udviklede sig i sidste ende til landplanter (Zhong et al., 2013; Delwiche og Cooper, 2015). Denne “terrestrialisering” af grønne planter repræsenterede en biologisk nøglebegivenhed, som for altid ændrede planetens biogeokemi og naturhistorie. Den efterfølgende udbredelse af landplanter ændrede atmosfæriske og yderligere ændrede jordbundsforhold og gjorde det muligt for andre forskelligartede livsformer at dukke op på land. Udviklingen af landplanter ændrede også menneskets historie mest markant gennem innovationen af landbruget og den deraf følgende skabelse af den moderne menneskelige civilisation. Delvis på grund af deres evolutionære betydning har charofytter fået betydelig opmærksomhed fra plantebiologer i løbet af de seneste årtier (Pickett-Heaps og Marchant, 1972; Pickett-Heaps, 1975; Mattox og Stewart, 1984; Becker og Marin, 2009; Harholt et al., 2016). Efterhånden som der er blevet indsamlet yderligere nye data om disse algers biologi, er de imidlertid blevet vigtige modeller til forståelse af grundlæggende fænomener inden for biokemi, cellebiologi, udviklingsbiologi, økologi og i stigende grad molekylærbiologi (Delwiche og Cooper, 2015; Lemieux et al., 2016). Frontiers in Plant Science-serien om “Charophytes” illustrerer disse organismers betydning for flere specifikke områder inden for plantebiologisk forskning. Denne mini-review fremhæver charofytternes egenskaber som modelorganismer inden for forskellige forskningsområder. Den vil forhåbentlig også opmuntre en ny generation af forskere til at udvide brugen af disse alger i grundforskning og til at indlede screening af andre taxa, som igen kan føre til identifikation og brug af nye modelcharofytter.
The “Line-Up” of Charophytes
I løbet af 1970’erne og 1980’erne blev ultrastrukturelle (f.eks, celledelingsmekanisme, flagellapparatets underbygning) og biokemiske (f.eks. enzymprofiler) data var de vigtigste kriterier for at inkludere en grønalge i charofyte-linjen (Mattox og Stewart, 1984; Lewis og McCourt, 2004; Leliaert et al., 2012). Fra 1990’erne til i dag har undersøgelser med fokus på molekylære analyser af kloroplast- og kernegenomer og transkriptomer bekræftet og forfinet tidligere taxonomiske og fylogenetiske skemaer (Timme et al., 2012; Delwiche og Cooper, 2015). Dette er yderligere blevet suppleret af detaljerede biokemiske, immunologiske og cellebiologisk baserede analyser af charofytcellevægge og hormonbiosyntese- og signalveje (Popper og Fry, 2003; Popper, 2008; Sørensen et al., 2010, 2011, 2012; Zhang og van Duijn, 2014; Ju et al., 2015; O’Rourke et al., 2015). Eksisterende charofytter udviser en relativt lav diversitetsprocent i sammenligning med andre grønne algetaxa og omfatter 13 familier og 122 slægter (Becker og Marin, 2009; Leliaert et al., 2012). Den nuværende fylogenetiske opfattelse placerer charofyterne i seks klasser (figur 1; Delwiche og Cooper, 2015). Den basale klasse, Mesostigmatophyceae, er repræsenteret af en enkelt kendt slægt, Mesostigma. Denne alge er en encellet biflagellat med en unik asymmetrisk form (dvs. som en fladtrykt redningsbåd) og er dækket af lag af ornamentalt udformede skæl (Manton og Ettl, 1965; Becker et al., 1991; Domozych et al., 1991, 1992). Den anden klasse, Chlorokybophyceae, består også af en enkelt kendt type, Chlorokybus atmophyticus. Denne sjældne alge danner en sarkinoid pakke af celler, der er omgivet af et tykt gel-lignende dække (Rogers et al., 1980). Det skal bemærkes, at alternative fylogenier placerer Mesostigma og Chlorokybus som søsterlinjer, der sammen er søsterlinjer til alle andre Streptofytter (Rodríguez-Ezpeleta et al., 2007). I den seksklassede fylogeni består den tredje klasse, Klebsormidiophyceae, af tre slægter, der danner enkle ugrenede filamenter (Sluiman et al., 2008), som ofte er medlemmer af “biologiske skorper”, der vokser på overflader i forskellige terrestriske habitater. Flere arter i denne klasse kan endda modstå betydelig udtørringsstress, når de indgår i ørkenskorper (Mikhailyuk et al., 2008, 2014; Holzinger og Karsten, 2013); andre er tolerante over for udtørringsstress, der er forbundet med miljøer med lave temperaturer (Stamenkovic et al., 2014; Herburger og Holzinger, 2015). Disse tre første klasser udgør de “tidligt divergerende” charofytter. Den “sene divergerende” lineage omfatter også tre klasser. Charophyceae, eller stenorme, findes almindeligvis i ferskvandsøkosystemer og besidder makroskopiske flercellede thalli bestående af aggregationer af forgrenede filamenter lavet af usædvanligt langstrakte celler (Lewis og McCourt, 2004). Medlemmer af denne gruppe udviser oogamy-baseret seksuel reproduktion, der omfatter bevægelige sædceller og ikke-motile æg, der begge produceres i flercellede gametangier. Coleochaetophyceae består af taxa, der har flercellede thalli lavet af forgrenede filamenter eller parenchyma-lignende thalli (Graham, 1993; Delwiche et al., 2002). Disse organismer udviser også en tydelig oogamy-baseret seksuel reproduktion. Coleochaetophyceae findes typisk i grænseområdet mellem ferskvand og terrestriske habitater, ofte som epifytter. Endelig består den største og mest forskelligartede gruppe af charofytter, Zygnematophyceae, af uniceller og ugrenede filamenter (Gontcharov et al., 2003; Guiry, 2013). Det kendetegnende for denne gruppe er tilstedeværelsen af konjugationsbaseret seksuel reproduktion, der kræver kompleks celle-celle-signalering og adhæsion (Abe et al., 2016). De zygnematofysiske alger er almindelige beboere af ferskvandshabitater, der undertiden forekommer i spektakulære efemeriske opblomstringer, samt i fugtige terrestriske substrater.

FIGUR 1. Eksisterende charofytter. Eksisterende Charophytes er opdelt i tidligt og sent divergerende taxa. De tidligt divergerende taxa omfatter Mesostigmatophyceae, Chlorokybophyceae og Klebsormidiophyceae, og de sent divergerende taxa omfatter Charophyceae, Coleochaetophyceae og Zygnematophyceae. Repræsentanter for hver gruppe er Mesostigma (Mesostigmatophyceae), Chlorokybus atmophyticus (Chlorokybophyceae), Klebsormidium flaccidum (Klebsormidiophyceae), Chara (Charophyceae), Coleochaete scutata (Coleochaetophyceae) og Micrasterias (Zygnematophyceae).
Charofytter som modelorganismer
En række charofytter er blevet anvendt i vid udstrækning som modelorganismer i studiet af grundlæggende biologiske processer. Deres små og enkle thalli (dvs. sammenlignet med landplanter) og deres lette eksperimentelle manipulation er blot to af deres egenskaber, der gør dem til attraktive modelorganismer. Nylige beviser har også vist, at mange charofytter har flere bemærkelsesværdigt lignende træk som landplanter, herunder tilstedeværelsen af biosyntetiske veje for mange vækstregulatorer (figur 2; Boot et al., 2012; Hori et al., 2014; Wang et al., 2014, 2015; Holzinger og Becker, 2015; Ju et al., 2015) og flere cellevægpolymerer (Popper, 2008; Sørensen et al., 2010, 2011, 2012; Mikkelsen et al., 2014). Disse to egenskaber har gjort charofytter effektive inden for sådanne studieområder som molekylær udvikling af planter og stressfysiologi. Mens mange charofytter er blevet brugt i en bred vifte af biologiske undersøgelser, er følgende taxa mest bemærkelsesværdige for deres omfattende brug som modeller inden for flere studieområder:
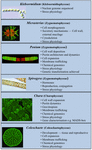
FIGUR 2. Nuværende model Charophytes. Flere charofytter, herunder Klebsormidium, Micrasterias, Penium, Spirogyra, Chara og Coleochaete, anvendes som modeller; hver af dem har specifikke egenskaber, der gør dem særligt velegnede som modeller for specifikke fysiologiske og biokemiske processer.
Desmider: De symmetriske modeller
Den Zygnematophyceae er for nylig blevet vist, at de højst sandsynligt er de nærmeste levende forfædre til landplanter (Wodniok et al, 2011; Delwiche og Cooper, 2015). Unicellulære medlemmer af den inkluderende placoderm desmidgruppe er blevet vigtige modeller til at belyse mange grundlæggende principper for plantecellebiologi og udvikling. Stor cellestørrelse, bemærkelsesværdig symmetri/form, tydelig cellevægsarkitektur og udførlige endomembran-/cytoskeletsystemer er blot nogle få af de karakteristika, der gør dem til fremragende cellesystemer til undersøgelse af cellefysiologi og udvikling. Den nylige etablering af stabile transformerede linjer og de snart sekventerede genomer/transkriptomer af udvalgte desmider har yderligere øget deres værdi i botaniske undersøgelser. To taxa, Micrasterias og Penium, har vist sig at være de mest velundersøgte af desmiderne.
Micrasterias har været den mest populære desmid til cellebiologisk forskning i de seneste 50 år på grund af dens unikke egenskaber (Meindl, 1993; Lutz-Meindl, 2016). Micrasterias-cellen udviser en bilateral symmetri, der ofte fremhæves af en spektakulært dissekeret (dvs. multilobed) periferi. Nye datterhalvceller, der produceres ved celledeling, har ikke denne komplekse morfologi, men er snarere sfæriske i form. Et komplekst post-cytokinetisk udviklingsprogram, der anvender flere sæt stærkt koordinerede subcellulære mekanismer, genererer efterfølgende den multilobede fænotype under dattersemicellens ekspansion. Dette program er også ansvarligt for aflejring af polymerer til fremstilling af både en primær og sekundær cellevæg. Disse begivenheder er centreret omkring et stort netværk af Golgi-legemer og tilhørende vesikler, der giver en kompleks sekretorisk mekanisme. Denne proces er til gengæld rettet mod specifikke celleoverfladesteder, der genererer en multipolær celleudvidelse og den samtidige sekretion af cellevægmakromolekyler og ekstracellulær slim (Kim et al., 1996; Lutz-Meindl og Brosch-Salomon, 2000; Oertel et al., 2004; Aichinger og Lutz-Meindl, 2005). Leveringen af sekretoriske komponenter til præcise celleoverfladelokaliteter kræver et udførligt actinomyosinbaseret cytomotilsystem og styres af flere signaltransduktionskaskader (Meindl et al., 1994; Oertel et al., 2003). Desuden er cellemorfogenese meget følsom over for ekstern stress (f.eks. oxidativ stress, saltholdighed, tungmetaller), der fører til store ændringer i celledifferentieringen (Darehshouri et al., 2008; Affenzeller et al., 2009; Andosch et al., 2012). Micrasterias er en fremragende organisme til analyse af disse subcellulære egenskaber, da den er let at vedligeholde og manipulere i laboratoriet, og den egner sig særligt godt til erhvervelse af højopløsningsbilleder ved hjælp af lys- og elektronmikroskopi, herunder immunocytokemi, FIB-SEM (Focused Ion Beam-Scanning Electron Microscopy) og EELS (Electron Energy Loss Spectroscopy) billeddannelse (Lutz-Meindl, 2007; Eder og Lutz-Meindl, 2008, 2010; Wanner et al., 2013; Lutz-Meindl et al., 2015). Desuden har indledende molekylære analyser, herunder produktion af transformerede cellelinjer, yderligere forbedret brugen af Micrasterias, især i den molekylære dynamik af cellevægsbehandling (Vannerum et al., 2010, 2011, 2012).
I løbet af det seneste årti er desmiden, Penium margaritaceum, også blevet en værdifuld modelorganisme (Domozych et al., 2009, 2014; Sørensen et al., 2014; Rydahl et al., 2015; Worden et al., 2015). I modsætning til Micrasterias har Penium en simpel cylindrisk form, besidder kun en primær cellevæg og aflejrer vægpolymerer på to specifikke steder på celleoverfladen under ekspansion (Domozych et al., 2011). Denne relative enkelhed er meget attraktiv for at belyse grundlæggende principper for plantecelleudvikling, herunder cellevægsudvikling, celleekspansion og sekretionsdynamik (Domozych et al., 2005, 2014; Ochs et al., 2014). En af dens vigtigste egenskaber er, at den har vægpolymerer svarende til dem, der findes i mange landplanter (f.eks. cellulose, pektiner, hemicelluloser), og at disse polymerer kan mærkes med monoklonale antistoffer. Efter mærkning af levende celler kan disse returneres til kulturen, hvor efterfølgende celleudvidelse og vægaflejringshændelser kan overvåges (Domozych et al., 2009; Rydahl et al., 2015). Penium er også let at vedligeholde i laboratoriet, og dens hurtige væksthastighed under præcist kontrollerede forhold gør den til en fremragende prøve til storskala samtidige mikroarray-screeninger af mange kemiske stoffer (ved vækst i multi-well-plader) og til vurdering af deres specifikke virkninger på ekspansion/vægudvikling (Worden et al., 2015). Dette hjælper betydeligt til at afsløre den rolle, som specifikke subcellulære komponenter og processer spiller i ekspansion/differentiering. For nylig har isolering af stabile transformerede linjer (Sørensen et al., 2014) yderligere øget potentialet af denne alge i fremtidige plantecelleundersøgelser.
Chara og Nitella: Cellulære giganter for plantefysiologi
Species af Chara og Nitella (Charophyceae) er længe blevet brugt som eksemplarer til en række biologiske undersøgelser, især dem, der beskæftiger sig med celledynamik, ekspansion og cytoplasmatisk streaming (Green, 1954; Probine og Preston, 1962). Deres makroskopiske thalli er kendetegnet ved knuder, hvor grene og gametangier opstår, og internodale regioner, der består af usædvanligt langstrakte celler. Internodale celler udviser en klart bemærkelsesværdig stratificering af subcellulære komponenter, hvorved spiralformede kloroplaster definerer en stationær cortex, der omgiver et endoplasma, hvor den hurtigst registrerede actinomyosin-genererede cytoplasmatiske strømning finder sted. Cytoplasmaet indeholder forskellige endomembrankomponenter, der deltager i dynamiske membrantrafikkenetværk (f.eks. endocytose og exocytose), der styres af komplekse signaltransduktionskaskader (Foissner og Wastenys, 2000, 2012, 2014; Sommer et al., 2015; Foissner et al., 2015; Foissner et al., 2016). De internodale celler besidder cellevægge, der er rige på cellulose og pektin, homogalacturonan (HG). Turgor er den vigtigste drivkraft for ekspansion, og pektin er sandsynligvis den belastningsbærende komponent, der styrer vægudvidelsen. Cyklisk, ikke-enzymatisk modulering og samtidig kalciumkompleksering af HG er den mekanisme, der er ansvarlig for at kontrollere vægmodulationer, der fører til celleekspansion (Proseus og Boyer, 2007, 2008, 2012; Boyer, 2016). En af de tekniske fordele ved Chara og Nitella i celleforskning er, at internodale celler kan fjernes individuelt; endoplasma og cellevægge kan “dissekeres” og derefter bruges som acellulære systemer til eksperimentel manipulation.
Andre thalluskomponenter som f.eks. rhizoider har organiseret polær organisering af cytoplasmaet. Disse celler indeholder sedimenterbare, mineralrige statolitter placeret ved spidsen, der fungerer i tyngdeopfattelse (Hodick et al., 1998; Braun, 2002; Braun og Limbach, 2006). Denne egenskab har gjort Chara til den første charophyte i rummet. Under TEXUS-raketens paraboliske flyvninger blev det vist, at statolitterne udøver spændingskræfter på aktinfilamenter. En balance af kræfter (dvs. tyngdekraften og den modvirkende kraft fra aktinfilamenterne) er ansvarlig for den korrekte placering af statolitterne i rhizoidet, som igen garanterer evnen til at reagere på tyngdevektoren (Braun og Limbach, 2006). Brugen af laserpincetter og langsomt roterende centrifuge-mikroskopi har vist, at statolit-sedimentation ikke er tilstrækkelig til at forårsage gravitropisk bøjning af rhizoidspidsen. De mineralrige statolitter skal snarere sætte sig på specifikke områder af plasmamembranen, for at de gravitropiske morfologiske virkninger kan fortsætte (Braun, 2002). Chara er også blevet brugt til at studere andre fænomener, herunder elektrofysiologi og hormoners rolle og bevægelse (Belby, 2016).
Coleochaete til udviklingsstudier og mønsterudvikling
Dannelsen af et flercelligt thallus (dvs. væv, organ) i en plante kræver præcise, rumligt regulerede celledelingsplaner, der styres af flere sæt af gener, der udtrykkes på specifikke punkter i løbet af udviklingen (Besson og Dumais, 2011; Umen, 2014). Thallus-morfogenese, der er styret i både 3-dimensionelt rum og tid, moduleres også som reaktion på eksterne stressfaktorer. I planter gøres disse udviklingsprogrammer endnu mere komplekse på grund af tilstedeværelsen af cellevægge, der ikke tillader cellevandring eller vævsfleksibilitet. Det er ofte vanskeligt at klarlægge de specifikke begivenheder, der er centrale for manifestationen af thallusens form og størrelse, i landplanter, hvor opløsningen af specifikke celleadfærd ofte er dårligt opløst, når de er indlejret i meget komplekse og ekspansive væv/organer. Det har været en fordel at anvende organismer med enklere thallusdesign til at forstå grundlæggende udviklingsfænomener, og charophyte-slægten Coleochaete er et sådant eksempel. Visse arter (f.eks. C. orbicularis, C. scutatum) producerer en parenchymatøs discoid thallus, der vokser udad ved en kombination af anti- eller peri-klinale celledelinger og efterfølgende ekspansion af de yderste celler (Brown et al., 1994; Cook, 2004). Delingsplanet for enhver celle følger enkle regler, der er baseret på cellestørrelse, celleform og geografisk placering i thallus (Dupuy et al., 2010; Besson og Dumais, 2011). Denne egenskab gør det muligt at opbygge matematiske modeller, som derefter kan bruges til at fortolke flercellet morfogenese (Domozych og Domozych, 2014; Umen, 2014) og give kritisk indsigt i organers og hele organismers evolution, biomekanik og fysiologi. Desuden er nogle Coleochaete-arter blevet undersøgt indgående for at bestemme de strukturelle og funktionelle modulationer, der sker i en thallus under tørkeforhold (Graham et al., 2012), dvs. nøgleelementer i forståelsen af tidlig landplanteevolution og plantevækstdynamik under tørke.
Insigt fra genom- og genstudier
Som mange andre teknologier har molekylære værktøjer evnen til at bibringe nye oplysninger af betydning for mange forskningsområder, herunder fysiologi og udvikling. F.eks. er medlemmer af MADS-box-genfamilien blevet isoleret og karakteriseret fra tre charofytter, Chara globularis, Coleochaete scutata og Closterium peracerosum-strigosum-littorale (Tanabe et al., 2005). Udtryksmønstret for MADS-box-generne i charofytterne tyder på, at de spiller en rolle i haploid udvikling og reproduktion (Tanabe et al., 2005). Det antages, at de er blevet rekrutteret til den diploide generation, og at deres roller i udviklingen er blevet yderligere diversificeret i løbet af udviklingen af landplanter (Tanabe et al., 2005). Til dato er Klebsormidium flaccidum den eneste charofyt, for hvilken der foreligger et næsten komplet kernegenom (Hori et al., 2014); selv om kloroplast- og mitokondrielle genomer er blevet sekventeret for repræsentanter for hver af de seks klasser af charofytter, og EST’er (expressed sequence tags) er tilgængelige for nogle charofytter (Delwiche, 2016). Klebsormidium-genomet har allerede afsløret tilstedeværelsen af gener til syntese af flere plantehormoner og signalformidlingsintermediater, mekanismer til beskyttelse mod høj lysintensitet (Hori et al., 2014) og tilstedeværelsen af gruppe IIb WRKY-transkriptionsfaktorer, som tidligere blev anset for først at være dukket op hos mosser (Rinerson et al., 2015). Flere charofyte-genomsekventeringsprojekter er i øjeblikket i gang (Delwiche, 2016) og vil muliggøre mere detaljerede komparative analyser, herunder et større antal charofyter, og potentielt give ny indsigt i de evolutionære relationer mellem arter og gener. Charofytters unikke position som den tidligst divergerende gruppe af Streptofytter gør dem til et særligt kraftfuldt redskab til evolutionært-udviklingsmæssige studier og nye teknologier som f.eks. den nylige opnåelse af stabil transformation i Penium (Sørensen et al, 2014) vil gøre det muligt at udforske funktionen af specifikke gener.
Fremtidige anvendelser af charofytter som modelorganismer
Et afgørende næste skridt i charofytforskningen er analysen af genomerne af inkluderende taxa og sammenlignende undersøgelser med dem af landplanter. Dette arbejde vil ikke blot give vigtig indsigt i de adaptive mekanismer, som charofytter udviklede, da de kom frem og med succes koloniserede terrestriske habitater, men vil også yderligere forfine og udvide brugen af de nuværende modelorganismer i den grundlæggende planteforskning. Selv om molekylære undersøgelser vil støtte mange forskningsområder, vil to områder, der umiddelbart vil være til gavn, være cellevægsbiologi og dynamikken af vækstregulatorer i vækst og udvikling. På førstnævnte område vil karbofytterne kunne bidrage til at klarlægge mange grundlæggende, men endnu dårligt opklarede fænomener, herunder pektin- og cellevægsproteinbiosyntese, den kontrollerede sekretion, deponering og postsekretoriske modulation af vægpolymerer og de specifikke, interaktive netværk for membrantrafikken i planteceller. Med hensyn til sidstnævnte vil karbofytmodellerne hjælpe med at tyde den molekylære signalering, cellebiologi og udviklingsdynamikken i forbindelse med vækst og udvikling, der er forbundet med hormoner som ethylen og auxin. Nylige økofysiologiske undersøgelser med charofytter tilbyder også en potentiel guldgrube af kritiske data til bestemmelse af, hvordan planteceller tilpasser sig stress, herunder udtørring og salttolerance (Holzinger og Karsten, 2013; Pichrtová et al., 2014; Herburger et al., 2015; Holzinger og Becker, 2015; Holzinger og Pichrtová, 2016; Kondo et al., 2016). Fremtiden for charofytforskning er virkelig meget lys og vil følgelig blive en velsignelse for hele plantebiologien.
Author Contributions
Alle forfattere på listen har ydet et væsentligt, direkte og intellektuelt bidrag til arbejdet og har godkendt det til offentliggørelse.
Interessekonflikt erklæring
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller finansielle relationer, der kunne opfattes som en potentiel interessekonflikt.
Akkreditering
Dette arbejde blev støttet af NSF (USA) MCB-samarbejdsbevillinger 1517345 og 1517546.
Belby, M. J. (2016). Eksperimentelt system til multiskala-salamander: fra elektrofysiologi af membrantransportører til celle-til-celle-forbindelse, cytoplasmatisk streaming og auxinmetabolisme. Front. Plant Sci. 7:1052. doi: 10.3389/fpls.2016.01052
CrossRef Full Text | Google Scholar
Braun, M. (2002). Tyngdeopfattelse kræver statolitter, der er aflejret på specifikke plasmamembranområder i rhizoider og protonemata fra salamandre. Protoplasma 219, 150-159. doi: 10.1007/s00709090200016
CrossRef Full Text | Google Scholar
Braun, M., og Limbach, C. (2006). Rhizoider og protonemata af salamanderalger: modelceller til forskning i polariseret vækst og plantegravitationssansning. Protoplasma 229, 133-142. doi: 10.1007/s00709-006-0208-9
CrossRef Full Text | Google Scholar
Graham, L. E. (1993). Oprindelse af landplanter. New York, NY: John Wiley & Sons, 287.
Google Scholar
Herburger, K., og Holzinger, A. (2015). Lokalisering og kvantificering af callose i Streptophyte grønalger Zygnema og Klebsormidium: korrelation med udtørringstolerance. Plant Cell Physiol. 56, 2259-2270. doi: 10.1093/pcp/pcv139
CrossRef Full Text | Google Scholar
Lemieux, C., Otis, C., og Turmel, M. (2016). Sammenlignende chloroplastgenomanalyser af streptofyte grønalger afdækker store strukturelle ændringer i Klebsormidiophyceae, Coleochaetophyceae og Zygnematophyceae. Front. Plant Sci. 7:697. doi: 10.3389/fpls.2016.00697
CrossRef Full Text | Google Scholar
Lutz-Meindl, U. (2007). Anvendelse af energifiltrerende transmissionselektronmikroskopi til billedgenerering og elementanalyse i planteorganismer. Micron 38, 181-196. doi: 10.1016/j.micron.2006.03.017
CrossRef Full Text | Google Scholar
Meindl, U. (1993). Micrasteriasceller som et modelsystem til forskning i morfogenese. Microbiol. Rev. 57, 415-433.
Google Scholar
Pichrtová, M., Kulichova, J., og Holzinger, A. (2014). Kvælstofbegrænsning og langsom tørring inducerer udtørringstolerance i konjugerende grønalger (Zygnematophyceae, Streptophyta) fra polære habitater. PLoS ONE 9:e113137. doi: 10.1371/journal.pone.0113137
CrossRef Full Text | Google Scholar
Pickett-Heaps, J. D., og Marchant, H. J. (1972). Fylogeni af grønalger: et nyt forslag. Cytobios 6, 255-264.
Google Scholar
Stamenkovic, M., Woelken, E., og Hanelt, D. (2014). Ultrastruktur af Cosmarium-stammer (Zygnematophyceae, Streptiophyta) indsamlet fra forskellige geografiske steder viser artsspecifikke forskelle både ved optimale og stress-temperaturer. Protoplasma 251, 1491-1509.
Google Scholar