
















































Denne illustration kan udskrives som sort/hvid farvelægningsark. Bareklik her og højreklik på billedet, der åbnes i et nyt vindue, og gem det på din computer.
Navn: Navn: C.megalodon(Stor tand)
Fonetisk: Meg-ah-low-don.
Navnet af: Meg-ah-low-don: Louis Agassiz – 1843.
Synonymer:: Calalodon: Navn: Procarcharodon megalodon,Megaselachus megalodon.
Klassifikation:: Forfatter: Agassigias, Hassami, Agassias, 1981: Chordata, Chondrichthyes,Elasmobranchii, Lamniformes.
Arter: Carcharodon megalodon,Carcharocles megalodon, Procarcharodonmegalodon eller endda Otodus megalodon, se teksten for en udførlig forklaring.
Diet: Kødædende.
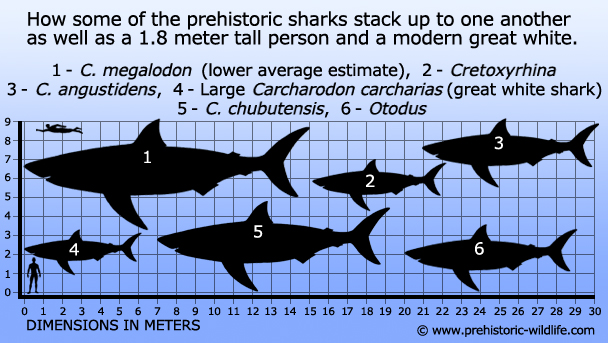
Størrelse: Der findes mange skøn, men normalt er den mellem 15 og 17 meter lang.
Kendte steder:
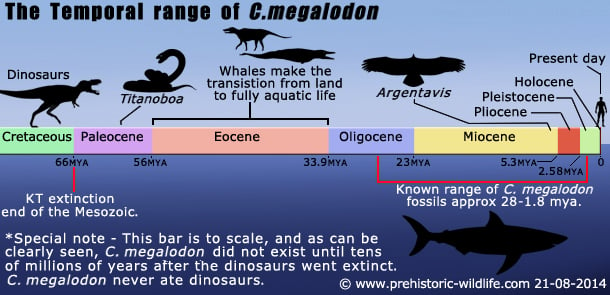
Orte: Overalt i verden: Tid: Sent oligocæn til tidligt pleistocæn.
Fossil repræsentation: Sent oligocæn til tidligt pleistocæn: For det meste tænder, men nogle ryghvirvler er også kendt.
Tænder og ryghvirvler – Fossiler af megalodon
Tænder er langt de mest almindelige rester af C. megalodon, og de større tænder nærmer sig 18 centimeter i højden. Det er usikkert, hvor længe folk har indsamlet C. megalodontænder, men det var først i 1667, at videnskaben anerkendte dem som hajtænder. Før dette tidspunkt troede folk generelt og ærligt, at C. megalodon var en haj.megalodontænder i virkeligheden var tunger fra drager, der var blevet forstenet (forstenet). Sandheden om dem kom frem, da den danske naturforsker Nicolas Steno korrekt identificerede en af disse “dragetænder” og omtalte den i sin bog The Head of a SharkDissected.
C.megalodontænder sammenlignes normalt med den store hvide hajs tænder på grund af deres overfladiske ligheder, nemlig at de er trekantede og savtakkede.På grund af denne lighed, især med tandsnit, mener man, at C. megalodon havde samme bidstil som den store hvide haj. Det ville indebære, at den bed ned i sit bytte og derefter rystede hovedet fra side til side, så tænderne savede sig gennem kødet.
Der er endda blevet fundet noget, der er tæt på at være komplette sæt C.megalodontænder fra samme individ, sammen.Disse afslører, at C.megalodon lignede andre hajer, idet de største tænder var forrest i munden. Jo længere fremme tænderne var, jo mere jævnt trekantede blev de. Jo mere sidelæns tænderne gik bagud i kæberne, jo mere indadvendigt buede blev de.Disse tænder hakkede sig fast i byttets kød og gjorde det sværere for byttet at undslippe, mens fortænderne skar, mens fortænderne skar.
En af de mest almindelige myter, der oftest fortælles om C. megalodon, er, at den kun kendes fra tænder. I virkeligheden er tænderne langt de mest almindelige fossiler af C. megalodon, men der er også fundet et lille og voksende antal hvirveldyr. I hajbiologien generelt består skelettet altid af “blødt” brusk, men hvirveldyrene består af hårdere forkalkede brusk.Det betyder, at mens hvirvlerne kan og ofte nedbrydes, kan de overleve længere tid, hvilket øger chancen for, at de bliver fossiliseret.

Størrelse -Optimering af den øvre størrelse af megalodon
Da tænderne fra C. megalodon blev erkendt, at det var tænder fra hajer, spurgte de fleste straks: “Men hvor stor var hajen?” Det ærligste svar er, at vi ikke helt ved det med sikkerhed, fordi der ikke er noget komplet eksemplar, som man kan tage et målebånd på.I stedet må palæontologer og iktyologer arbejde med det, de har, i dette tilfælde tænderne. Der har været mange undersøgelser af hajens længde baseret på analyse af tænder, hvoraf de fire mest almindeligt omtalte er her.
En af de metoder, der engang blev brugt til at måle længden af C. megalodon, var at måle højden af tandemaljen. Denne metode blev udviklet af John E.Randall i 1973 og resulterede i et skøn på 13 m. Metoden anses dog ikke for nøjagtig af det bredere videnskabelige samfund, da metoden først blev udviklet ved at sammenligne tænderne fra den store hvide haj, og selv om disse tænder umiddelbart ligner hinanden, er de faktisk helt forskellige fra C. megalodons tænder.Desuden kan mængden af emalje på en tand variere selv blandt tænder af samme art på grund af slid og dårlig bevarelse.
En anden vurderingsmetode, der blev udviklet i 1996 af Michael D. Gottfried, Leonard J. V. Compagno og S. Curtis Bowman, fungerer på grundlag af en korrelation mellem den skrå højde af en tand og hajens længde.Skrå højde er ikke den samlede højde, men længden fra tandspidsen til den laterale kant og er tandens faktiske skærelængde. Igen var denne undersøgelse baseret på en undersøgelse af store hvide tænder, og den gav et skøn på 15,9 meter. Men selv om denne metode er pålidelig, kan den kun give skøn ud fra de givne tænder og ikke nødvendigvis ud fra arten.
I2002 gav dr.Clifford Jeremiah opstillede en anden metode baseret på bredden af en tandrod, den del under kronen, der er dækket af kød. Dette er et simpelt princip, som går ud på, at bredden af tandkronen kan bruges til at beregne bredden af kæben, som igen kan bruges til at beregne længden af hajen. I de enkleste vendinger fungerer den på grundlag af, at der er 1 centimeter tandrod for ca. hver 129 centimeter kropslængde.Dette resulterede i et skøn over længden på 15,5 meter for en tand, der havde omkring 12 centimeter rod.
I 2002 udviklede Dr. Kenshu Shimada en anden mere kompleks metode baseret på tandkronens proportioner. Da denne metode blev anvendt på den tand, som Gottfried et al. havde anvendt i 1996, kom den anslåede længde frem til 15,1 meter.Det største skøn, som denne metode nogensinde har givet, blev opnået på en særlig stor tand fra Panama, som gav en kropslængde på 16,8 meter.
At gå med en anslået kropslængde i intervallet 15-16 meter anses for at være ret sikkert for C. megalodon, idet høje skøn, der nærmer sig 17 meter, anses for at være realistisk mulige.Der findes større skøn med påstande om, at C. megalodon kunne nærme sig og endda overskride en længde på 20 meter, men de fleste af disse skøn anses kun for at være muligheder og afspejler ikke de nuværende fossile beviser. Selv med en mere beskeden længde på 15 meter ville C. megalodon stadig have overgået enhver anden haj, enten levende eller uddød, og selv en stor hvid haj ville have blegnet i sammenligning med dens gigantiske størrelse.
Hvor meget C. megalodon er større end C. megalodon, er der flere skøn. megalodon vejede, er et andet spørgsmål, der skal undersøges, selv om dens enhedsvægt kan give et endnu bredere spektrum af skøn. De nuværende skøn er igen baseret på sammenligning med den store hvide haj, hvor et skøn for en 15,9 meter C. megalodon er vurderet til 47 tons. Større skøn omfatter 59 tons for en 17 meter C. megalodon og 103 tons for en 20,3 meter C. megalodon.Som du måske har bemærket, stiger vægten af en C. megalodon ikke med et konstant beløb med længden, hvilket er grunden til, at en 20,3 meter lang haj er omkring en tredjedel længere end en 15,9 meter lang haj, men alligevel vejer mere end det dobbelte af det lidt mindre eksemplar. Dette skyldes, at man skal være klar over, at hajen ikke blot er længere, den er også forholdsmæssigt tykkere og mere omfangsrig med øget muskelmasse til at bære den større ramme. Det betyder også, at et dyr, hvad enten det er en haj eller en anden art, altid vil nå en størrelse, hvorfra det ikke kan vokse yderligere på grund af, hvad dets levested kan bære af føde, hvilket er en naturlig kendsgerning baseret på den logik, at en større krop har brug for et større indtag af føde for at få energi til at drive den.
Måske Biologi – Det indre arbejde i megalodon
På grund af dens store kropsstørrelse C. megalodon menes det, at den næsten helt sikkert har levet med virkningerne af gigantotermi. Dette er, når et dyr er så massivt, at dets krop holder på varmen, idet de ydre lag af muskler og kød faktisk isolerer de indre organer fra omgivelserne. Dette forårsager en stigning i stofskiftet, hvilket igen gør dyret mere aktivt. C. megalodon kan have taget processen med gigantotermi endnu længere ved at lede strømmen af varmere blod ind i hovedet og snuden, som den store hvidhaj er kendt for at gøre. Dette ville have den virkning, at C. megalodons hjerne og sanseorganer som f.eks. lugtesansen, synet og de elektroreceptive ampuller fungerer med varmblodet stofskifte.Det faktum, at C. megalodons tænder er kendt fra alle kontinenter, er bevis på, at den havde en kosmopolitisk udbredelse ligesom mange andre succesfulde havrovdyr.Men udbredelsen af C. megalodon synes at være blevet reduceret, efterhånden som havene blev afkølet, især mod slutningen af Pliocæn.
Den samlede fremtoning af C. megalodon ville sandsynligvis have været meget lig de lamniforme hajer, der svømmer i havet i dag, selv om dens store størrelse og iboende kraft betyder, at den sandsynligvis var ret kraftig i forhold til moderne hajer.Bruskskelettet ville sandsynligvis også have været mere robust for at kunne klare de større belastninger fra større muskler og kraftigere byttedyr.
C.megalodon havde sandsynligvis en bevægelig overkæbe, der ikke kun kunne bevæge sig op og ned, men også kunne projiceres fremad uafhængigt af resten af kroppen.
Denne slags kæbe kan ses hos moderne hajer i dag og er grunden til, at en haj, der er ved at bide sig fast i noget, ser meget anderledes ud end en haj, der bare sejler rundt.Når overkæben projiceres fremad, bliver overkæben og tænderne faktisk synlige, mens de normalt er skjult af munden. Med en kæbe, der bevæger sig på denne måde, ville C. megalodon gribe fat i sit bytte med sin langsommere kæbe, der holdt det på plads, og derefter ryste hovedet fra side til side, så tænderne skar et stykke kød af.Overkæben trækkes derefter tilbage, hvilket har den virkning, at den trækker kødstykket ind i munden. Hajen gentager derefter denne proces, indtil den har spist sig mæt.
Et studieområde, der er umiddelbart forbundet med opdagelsen af tænderne, er vurderingen af den bidekraft, som C. megalodon var i stand til. Dette er en vigtig ting at vide, da det, hvor kraftigt et dyr kan bide ned på noget, kan føre til ledetråde til, hvilken slags bytte et dyr spiste og hvordan det spiste det.I 2008 blev der skabt en biomekanisk computermodel til at vurdere bidekraften hos store hvide hajer. Da denne model blev anvendt på C. megalodon, viste det sig, at en 15,9 meter lang C. megalodon, et stort gennemsnit, var i stand til at udøve en bidekraft på 108 514 newton, lidt over 11 tons.Når bidekraften for en C. megalodon på 20,3 meter, den øverste ende af den estimerede potentielle C. megalodon-størrelsesskala, blev anslået, var resultatet 182 201 newton, over 18,5 tons. Dette betyder, at selv de lavere estimater for C. megalodon er meget store. megalodon stadig giver den en meget højere bidkraft end den glubske placoderm Dunkleosteus og endda højere end den mægtige Tyrannosaurusrex. En anden ting, der skal tages i betragtning, er, at C. megalodon sandsynligvis rystede sit hoved fra side til side ligesom andre hajer med et lignende gebis. Det betød, at de faktiske kræfter, som C. megalodons uheldige bytte blev udsat for, sandsynligvis var endnu større.
Hvorimod de fleste af undersøgelserne af C. megalodons megalodon er koncentreret om tænderne, er et område, der ofte overses, ryghvirvlerne. På grund af det lille antal kendte ryghvirvler er det ikke alle, der kan studere dem, som de kan studere tænderne, men en ting, som ryghvirvlerne tydeligt viser, er koncentriske ringe. Disse koncentriske ringe er i princippet de samme som de vækstringe, man kan se i en træstub, og de er også synlige hos andre moderne hajer, der svømmer i havene i dag.Optælling af disse koncentriske ringe har givet et skøn på 25 til 40 års alder for den C. megalodon, der er kendt fra fossiler, og palæontologer foreslår, at C. megalodon kan have været i stand til at leve endnu længere end dette.
Unge megalodon – unger og børnehaver
Med deres store størrelse og pelagiske levevis er C. megalodon en stor haj, der kan leve i endnu længere tid. megalodon menes at have født levende unger. Præcis hvordan C. megalodond gjorde det, er dog stadig omdiskuteret, da hajer har to metoder til at gøre det. Den første kaldes viviparitet, og det er her, hvor ungen, som er det, en babyhaj kaldes, vokser inde i moderen, indtil den er klar til at blive født. Hajunger, der fødes på denne måde, får næringsstoffer via en navlestreng, og når ungen er født, bliver moderkagen normalt skubbet ud lige bagefter.
Den anden metode er ovoviviparitet, og her udvikler hvalpen sig inde i et æg. Hos hajer, der udviser ovoviviparitet, lægges ægget dog ikke, men bevares inde i hajmoderen. Hajunger af disse hajer er ikke knyttet til deres mor med en navlestreng, men bruger i stedet æggeblommen i løbet af deres udvikling.I begge udviklingstyper er ungerne ved bevidsthed og i stand til at svømme ved egen kraft, når de passerer ud af moderens krop. C.megalodon-unger er normalt afbildet som kommer ud med halen først, og selv om dette anses for at være den sædvanlige metode, er nogle hajarter i dag faktisk født med hovedet først.
Som vi ser det hos hajer i dag, er C. megalodon sandsynligvis ikke fødte deres unger et hvilket som helst sted, men valgte i stedet det, der kaldes børnehaveområder. Et børnehaveområde er et sted, hvor en ung haj kan leve og jage, mens den er i sikkerhed for andre større rovdyr. Hajer i børnehaveområder betragtes som generalister, der angriber og spiser alle slags dyr, herunder fisk, blæksprutter (som blæksprutte og blæksprutte), skildpadder og stort set alt andet, de kan fange.
For at identificere potentielle opvækstområder leder palæontologer efter koncentrationer af mindre C. megalodon-tænder. Et område, der synes at være rigeligt med disse tænder, er Mellemamerika og de sydligste områder af USA. Hvis man går tilbage til Oligocæn, eksisterede Panama ikke, fordi det daværende høje havniveau lagde en stor del af området under vand. Dette område var kendt som den mellemamerikanske havvej og udgjorde en oceanisk passage mellem Stillehavet og Atlanterhavet.Området havde sandsynligvis et enormt område med lavt vand, der simpelthen ikke var dybt nok til, at større rovdyr kunne operere der, hvilket gjorde det forholdsvis sikkert for de meget mindre C. megalodonunger.

Fossile beviser tyder stærkt på, at der i løbet af Miocæn var en meget stor bestand af unge C. megalodonunger i de lavvandede farvande mellem Nord- og Sydamerika i en meget høj bestand af unge C. megalodonunger. megalodon, mens de større voksne dyr strejfede rundt i de dybere farvande i det åbne hav (rød).Fossile forekomster tyder også på, at tidlige hvaler som f.eks. hvaler (lilla) brugte det, der dengang var kendt som den mellemamerikanske havvej som en passage mellem Stillehavet og Atlanterhavet. Høj vulkansk aktivitet og faldende havniveauer lukkede denne passage, hvilket resulterede i en reduktion af tilgængelige byttedyr samt et dramatisk skift i havstrømmene. Oven til højre ses Mellemamerika, som vi kender det i dag, med de varme havstrømme.
Undersøgelse af de mindre tænder viser, at C. megalodon i opvækstområderne var så små som 2 til 3 meter lange. Det betyder dog ikke, at de var så store, da de blev født, men blot at tænderne kom fra unge individer, som var aktive i området.Efterhånden som de unge C. megalodon blev ældre, blev de også større, og som sådan måtte de til sidst forlade de lavvandede områder til fordel for det åbne havliv. Dette ville indlede den anden fase af deres liv, hvor de skulle specialisere sig i at angribe store havdyr.
Bytteobjekter – Hvad spiste megalodon?
En ting, der straks skal ryddes af vejen, er, at C. megalodond ikke spiste på dinosaurer. Dette er en myte, der spredes i populærkulturen, især i film og romaner, hvor C. megalodons “skurk” bliver bygget op for at være mere dramatisk. C. megalodond optræder ikke i de fossile optegnelser før slutningen af Oligocæn, ca. 36 millioner år efter, at dinosaurerne uddøde i slutningen af Kridttiden, hvilket gør det umuligt for C. megalodond at blive spist af dinosaurer. megalodon at have spist endinosaurer (for en haj, der virkelig kunne have spist dinosaurer og store marine krybdyr, se Cretoxyrhina).
Den foretrukne føde for C. megalodon synes at have været hvaler og især små til mellemstore hvaler. Der er også beviser for, at C. megalodon har spist hvaler.megalodon angreb og spiste store havskildpadder, som formentlig var for langsomme til at undslippe, selv om deres panser ikke beskyttede dem mod C. megalodons kolossale bidkraft. Men hvad C. megalodon jagede, var afhængig af individets alder, idet mindre C. megalodon jagede dyr som f.eks. gugong, og større ældre C. megalodon jagede de større hvaler.
C. megalodons angrebsstrategi var afhængig af individets alder, idet mindre C. megalodon jagede dyr som f.eks. megalodon var ikke ligefrem raffineret, men den var effektiv til at nedlægge hvaler. Nogle hvalers ryghvirvler viser kompressionsskader, som er blevet fortolket som værende forårsaget af et pludseligt og massivt nedslag nedefra. Dette gjorde det muligt at rekonstruere et scenario, hvor C. megalodon ville nærme sig en hval nedefra for at undgå at blive set af sit mål.Når den havde gjort sig klar til et angreb, ville C. megalodon bruge sine stærke muskler til med høj hastighed at bevæge sig op til overfladen og slå ind i hvalen nedefra. Hvis hvalen ikke endte i C. megalodons kæber, ville den højst sandsynligt være blevet bedøvet af slaget, hvilket gav C. megalodon tid til at foretage et dræbende bid.Der findes dog mindst én fossil ryghvirvel, som viser, at den blev udsat for denne form for angreb, men alligevel formåede at hele. Dette viser, at i dette tilfælde overlevede heldyret ikke blot angrebet, men levede også længe nok til, at skaderne kunne hele.

Særlig bemærkning* – Ovenstående sekvens har til formål at illustrere en mulig metode, som C. megalodon-hajer jagtede byttedyr på. Det er ikke hensigten at antyde, at dette var den eneste måde, som C. megalodonhajer jagede på.
Undersøgelse af fossiler, der synes at stamme fra C. megalodonbyttet, viser, at C. megalodon faktisk gik målrettet efter de knoglelignende områder som f.eks. brystkassen. Her havde C. megalodon to ting til sin fordel, ekstremt robuste tænder, der ikke gik let i stykker, og en knusende bidkraft, der let kunne knoglebryde knogler, hvilket igen forårsagede store skader på de indre organer, som de skulle beskytte.Yderligere støtte for denne metode kommer fra kompressionsbrud på tænderne, hvor de er blevet afstumpet, hvilket tyder på kraftige slag med et hårdt stof som knogler.
Når C. megalodon angreb større hvaler, der muligvis var for store til et bid i et område som f.eks. ribbenene, ændrede den sin taktik. I stedet for at gå efter de kritiske organer angreb C. megalodon halen for at forsøge at immobilisere sit bytte.Det er en meget intelligent strategi, for selv om hajer næsten hele tiden svømmer fremad, så de kan trække vejret, kan de kun opretholde ekstremt høje forfølgningshastigheder i korte perioder. Det skyldes, at hajens hvide muskel (ca. 90 % af den samlede muskelmasse), som bruges til at give pludselige fartbølger, bliver meget hurtigt træt, mens hajens røde muskel (ca. 10 % af den samlede muskelmasse) har mindre kraft, men en ufattelig stor udholdenhed, hvorfor den mindre røde muskel bruges til normal fart.Ved at forkrøblere en stor hval kunne C. megalodon tage sig god tid til at spise i stedet for at overanstrenge sig.
Nogle har hævdet, at C. megalodon simpelthen var for stor til at jage og kun kunne have været en ådselæder. I lyset af overvældende fossile beviser, der viser skader, ikke kun tandmærker, på mange store hvaler, anses en sådan påstand ikke bare for usandsynlig, men næsten umulig.Selv om de fleste hajer og kødædende dyr generelt vil benytte lejligheden til at spise fra et kadaver, gør det dem ikke udelukkende til ådselsædere. Havdyr, der kun lever af at æde, har også en tendens til at være bundædere, der venter på, at døde dyr synker ned på havets bund. Ved at anslå størrelsen af C.megalodon har man også anslået, hvor meget føde det ville kræve at holde den i live.Mængden varierer meget, men svinger mellem 600 og 1200 kg mad om dagen. Det er en enorm mængde mad for en ådselæder, da ådselædere er tilpasset til at kræve meget lidt energiudnyttelse, fordi de ikke ved, hvornår eller hvor deres næste måltid kommer fra. Når man tager alle fossile beviser, biometriske modeller og viden om hajernes levevis og biologi i betragtning, er resultatet, at ådselæder er den mindst sandsynlige overlevelsesmetode for C. megalodon.

Udslettelse – Hvorfor forsvandt megalodon?
C.megalodondis forsvinder fra de fossile optegnelser nær slutningen af den første fase (Gelasian) af Pleistocæn for 1,8 millioner år siden. Denne forsvinden er markeret ved den konstante nedgang i C. megalodondis’ antal. Megalodon-fossiler, indtil de forsvinder helt. Selv om der er nogle få teorier om, hvorfor C. megalodon forsvandt, er det mest sandsynligt, at det var en række skiftende begivenheder, der førte til dens undergang, snarere end blot én ting.
Den udløsende begivenhed for udryddelsen af C. megalodon synes at have været den globale nedkøling. Hvis C. megalodon havde et varmblodet stofskifte gennem gigantotermi, ville den til at begynde med have brug for et større kalorieindtag end et helt koldblodet væsen af samme størrelse. Jo koldere vandet er, jo større er forskellen, hvilket betyder, at C. megalodon ville have haft brug for en endnu større mængde føde for at klare temperaturfaldet. Desuden er gigantotermi stadig ikke nogen erstatning for et ægte varmblodet stofskifte, og en haj i koldere farvande ville stadig muligvis have været mere træg end i varmere farvande, hvilket yderligere ville hæmme dens evne til at jage.
En følgevirkning af de koldere globale temperaturer er, at store mængder vand begyndte at størkne til is, hvilket fremgår af tilstedeværelsen af store iskapper på hele den nordlige halvkugle.Tilstedeværelsen af mere is betød, at havniveauet faldt, og den mest dramatiske konsekvens heraf var dannelsen af Panamas isharmus, hvilket også blev hjulpet på vej af nye landformationer, der blev opbygget af den fortsatte vulkanske aktivitet i dette område.Dette skabte i det væsentlige en landbro mellem Nord- og Sydamerika og isolerede Stillehavet og Atlanterhavet fra hinanden på dette punkt.
Det umiddelbare resultat af dannelsen af Panamas isharmus var lukningen af den mellemamerikanske havvej, der tilsyneladende blev brugt som en vigtig vandringsrute for hvaler, som det fremgår af den store koncentration af hvalfossiler.Dette falder sammen med en generel nedgang i antallet af hvalarter, idet langt mindre end halvdelen af de pleistocæne hvaler har overlevet til nutiden. I dag findes der kun 6 hvalslægter i modsætning til over 20 slægter i Miocæn.De resterende hvaler var stadig vandrende, men syntes at have foretrukket de polare områder, formentlig på grund af den større mængde hvirvelløse føde, som Baleen-hvaler er tilpasset til at spise.Tandhvalerne synes heller ikke at have været en levedygtig mulighed, da deres antal også blev dramatisk reduceret, idet kaskelothvalen var den eneste store tandhval, der overlevede til i dag. Da C. megalodon var begrænset til de varmere havområder, havde den ikke længere konstant adgang året rundt til den fødeforsyning, som den var mest tilpasset til at dræbe.
Den store størrelse af C. megalodon arbejdede utvivlsomt imod den i disse tider, da de eneste andre tilgængelige byttedyr var mindre og hurtigere, og selv om de blev fanget, gav de ikke den samme næring som de større hvaler.Kannibalisme er også blevet foreslået som en mulig overlevelsesstrategi for C. megalodon, men det ville kun fungere, så længe der var andre C. megalodon at spise. Hvis dette faktisk skete, ville kannibalisme kun udtynde C. megalodonnumrene yderligere, hvilket igen ville begrænse det antal, der ville reproducere sig.
Det er forbundet med det potentielle tab af opvækstområder som følge af ændringer i havniveauet.Selve oprettelsen af Isthmus of Panama synes faktisk også at have fjernet et sådant opvækstområde, hvilket fremgår af et stort antal unge C. megalodontænder fra dette område.Et andet formodet opvækstområde var Maryland, som lå så langt mod nord, at vandet måske blev for koldt til at bære C. megalodon. Tabet af opvækstområder betyder, at C. megalodon-ungerne selv ville have været mere modtagelige over for rovdyr, måske endda andre C. megalodon, når de forsøgte at overleve.
Den sidste teori omfatter fremkomsten af nye rovdyr med særlige henvisninger til udviklingen af rovdyr af typen delfinider, som i dag repræsenteres af orkaen, også kendt som spækhuggerhvalen. Efterhånden som antallet af C. megalodon faldt, steg antallet af delfinider.Det er dog vanskeligt at sige, om disse nye rovdyrs fremkomst spillede en rolle i C. megalodons tilbagegang, da det også kunne være C. megalodons tilbagegang, der gav de nye rovdyr plads til at trives.Der er fossile beviser, der viser rovdyr/byttedyr-interaktion mellem C. megalodon og C. megalodon. megalodon og delfiniderne, som det fremgår af C.megalodontandmærker på delfinideknogler.
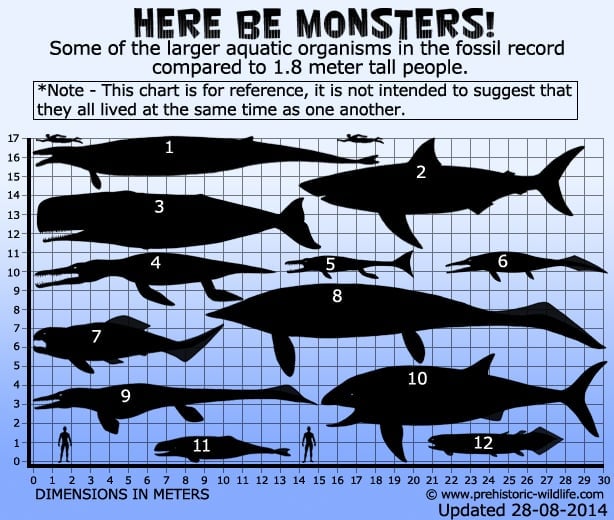
1 – Basilosaurus(hval), 2 – C.megalodon – lavere gennemsnitligt estimat (haj), 3 – Livyatanmelvillei – lavere estimat (hval), 4 – Pliosaurusfunkei, alias Pliosaurusfunkei, a.k.a Predator X (pliosaur), 5 -Plesiosuchus(thalattosuchian), 6 – Thalattoarchon(ichthyosaur), 7 – Dunkleosteus(arthrodire placoderm), 8 – Shastasaurus(ichthyosaur),9 – Tylosaurus(mosasaur), 10 – Leedsichthys- øvre estimat(fisk)), 11 – Brygmophyseter(hval), 12- Rhizodus(fisk med lobfinnede finner).
Sidste overlevende?
Nogle mener, at C. megalodon overlevede den pleistocæne æra og stadig svømmede i havene så sent som i den holocæne æra. Deres bevis for denne påstand kommer fra en delvis C. megalodon-tand, der blev opdaget i 1872 af besætningen på HMS Challenger, og som, da den blev testet i 1959, blev anset for kun at være 10.000 år gammel.Denne test målte imidlertid niveauet af mangandioxid på fossilet, en metode, der nu betragtes som mangelfuld på grund af de forskellige grader af mangandioxid, der kan ophobes på forskellige fossiler, selv fra samme æra. Da tanden blev underkastet senere radiokulstofdateringsteknikker, viste det sig, at den havde et for lavt kvælstofniveau til at kunne testes, og derfor er tanden siden blevet anset for at være uprøvelig, og tidligere skøn over, at C. megalodon uddøde i løbet af det tidlige pleistocæn, er fortsat gyldige.

Klassifikation – Er megalodon beslægtet med den store hvide haj?
Det største stridspunkt i forbindelse med C. megalodon er nok, om den faktisk er beslægtet med den store hvide haj, som vi kender i dag. Sammenligninger mellem C. megalodon og C. megalodon er ikke nødvendigvis en del af det samme. megalodon og den store hvide haj skyldes for det meste det princip, at C. megalodon var den største haj, og at den store hvide haj er den største haj, som vi kender i dag. Også tænderne mellem de to ses ofte som nogenlunde ens.Tilhængere af C. megalodon inden for Carcharodon peger på, at tandligheden er et resultat af, at både C. megalodon og den store hvide haj nedstammer fra Palæocarcharodonorientalis.
Problemet med at sammenligne C. megalodon med Carcharodon er, at C. megalodon og den store hvide haj er et resultat af, at C. megalodon og den store hvide haj nedstammer fra Palæocarcharodonorientalis. megalodon og den store hvide haj på grundlag af lignende tænder er, at de eneste ligheder, der findes, er, at begge hajer har tænder, der er trekantede og savtakkede. Ud over dette er den store hvide hajs tænder mere gracile, idet de er meget tyndere end C. megalodons tænder. Desuden er C. megalodon og C. megalodon meget tyndere, mens C.megalodon anses for at have et generelt lignende tandlayout som den store hvide haj, er den tredje fortand (tredje fortand fra midten af overkæben) hos C. megalodon anderledes, idet den peger nedad som de to første, hvilket er anderledes end hos den store hvide haj. C. megalodons fortænder har også et karakteristisk “ar” formet som en chevron, der ligger mellem tandens krone og rod, hvilket ikke er tilfældet hos den store hvide haj.
Et alternativ til at placere C. megalodon i slægten Carcharodonshark ville være at placere den i den ældre Carcharoclesgenus. Hovedargumentet for denne placering er, at en anden stor gammel haj ved navnCarcharoclesauriculatus faktisk menes at være en forfader til C. megalodon. Megalodon. Med tænder, der målte op til næsten 12 centimeter i længden, var Carcharoclesauriculatus stor, men i alt en tredjedel mindre end C. megalodon, hvis man sætter den i forhold til en 18 centimeter lang C. megalodon. Det er dog muligt, at Carcharoclesauriculatus kunne være vokset sig større og have givet anledning til C. megalodon, da rovdyr i alle miljøer har en tendens til at blive ved med at vokse sig større, indtil deres miljø ikke længere kan bære yderligere vækst.
En sådan placering af C. megalodon inden for Carcharocles ville faktisk afslutte en overgang, hvor hajerne mistede laterale hakker på deres tænder.Denne overgang begynder med de laterale tandhuler, der er tydeligt til stede i Otodusobliquus, de reducerede tandhuler i Carcharoclesauriculatus, til ingen tandhuler i C. megalodon.Disse tænder har ogsåchevronformede ar, hvor kronen møder roden, noget, der ikke findes i store hvide tænder.
En anden teori går ud på, at C. megalodon var forfader til den store hvide haj, og at hajen med tiden blot blev mindre. Det største problem med denne tankegang er imidlertid, at den store hvide haj faktisk svømmede i havet længe før C. megalodon blev født. megalodon blev uddød, idet fossiltænder af den store hvide haj dukkede op i midten af Miocæn for 16 millioner år siden, over 14 millioner år før C. megalodon uddøde. Tilhængere af teorien insisterer stadig på, at den store hvide haj kan have udviklet sig fra en mindre art af C. De, der kender til sabeltandkatten Smilodon, er måske klar over, at der var tre helt forskellige arter af samme slægt, som ikke blot synes at nedstamme fra den samme forfader, men som også i en periode var aktive sammen i samme tidsrum.Det er ikke så langt ude, at lignende hændelser kunne ske i andre dyregrupper. Alligevel synes der ikke at være noget overgangsled, der viser ændringen fra C. megalodon til den store hvideast – tænderne ville ikke bare blive mindre, de ville støt ændre sig til den store hvideform.Hvis den store hvidhaj er beslægtet, er det mere sandsynligt, at den store hvidhaj har en fælles umiddelbar forfader med C. megalodon.
I 2012 fik teorien om, at C. megalodon og Carcharodoncarcharias ikke er beslægtede, lidt mere støtte med beskrivelsen af en ny art af Carcharodon, Carcharodonhubbelli, også kendt som Hubbell’s hvidhaj. Tænderne hos Carcharodon hubbelli er af nogle blevet fortolket som overgangstænder, der forbinder Carcharodon carcharias med Isurusgenus, som huser makohajerne. Da de fleste forskere ikke mener, at der er en direkte forbindelse mellem makohajer og megatoothedshajer som C. megalodon, kan dette tyde på, at Carcharodoncarcharias faktisk er adskilt fra C. megalodon, og at C. megalodon i forlængelse heraf bør placeres inden for Carcharoclesgenus.
Som der er ligheder i den overordnede morfologi hos C. megalodon og den store hvidhaj, er det mest sandsynlige resultat af evolution snarere end af genetisk avl.Denne grundlæggende kropsform kaldes fusiform, eller mere løst “torpedoformet”, og er baseret på en spids forreste del, der stiger op til et bredt centrum, før den spidser til til et andet punkt bagud. Denne form har gentaget sig selv i naturen utallige gange og er bestemt ikke enestående for hajer, da det simpelthen er den mest effektive form til at bevæge sig under vandet. Nogle har også anset megalodon-hajen for at tilhøre enten Procarcharodon- eller Otodus-slægten, hvilket skyldes forskning, der tyder på en synlig overgang fra Otodus-slægtens tænder til megalodon-slægtens tænder, som levede senere. Igen er der dog stor forskel på forskerne i forhold til, hvilken slægt de megalodon-arter hører til.
Videre læsning
– Recherches sur les poissons fossiles/par Louis Agassiz – Neuchatel:Petitpierre. p. 41. – Louis Agassiz – 1833-1843.
– Size of the Great White Shark (Carcharodon) – Science Magazine 181(4095): 169-170 – John Randall – 1973.
– Carcharodon megalodon from the Upper Miocene ofDenmark, withcomments on elasmobranch tooth enameloid: coronoi’n – Bulletin of theGeological Society of Denmark (Copenhagen: Geologisk Museum) 32: 1-32.- Svend Erik Bendix-Almgreen – 1983.
– Catalogue of Cuban fossil Elasmobranchii (Paleocene to Pliocene) andpaleogeographic implications of their Lower to Middle Mioceneoccurrence – Boletín de la Sociedad Jamaicana de Geología (Cuba) 31:7-21 – M. Iturralde-Vinent, G. Hubbel & R. Rojas – 1996.
– The Megatooth shark, Carcharodon megalodon: Roughtoothed, hugetoothed – Mundo Marino Revista Internacional de Vida (non-refereed)(Marina) 5: 6-11. – J. C. Bruner – 1997.
– Fossile hajer fra Jamaica – Bulletin of the Mizunami Fossil Museum.pp. 211-215. – Stephen Donovan & Gunter Gavin – 2001.
– An associated specimen of Carcharodon angustidens(Chondrichthyes,Lamnidae) from the Late Oligocene of New Zealand, with comments onCarcharodon interrelationships – Journal ofVertebrate Paleontology 21(4): 730-739. – M. D. Gottfried & R. E. Fordyce – 2001.
– The relationship between the tooth size and total body length in thewhite shark, Carcharodon carcharias (Lamniformes:Lamnidae) – Journalof Fossil Research (Japan) 35 (2): 28-33. – Kenshu Shimada – 2002.
– Ny registrering af lamnidehajen Carcharodon megalodon fra det mellemste miocæn i Puerto Rico – Caribbean Journal of Science 39: 223-227. -Angel M. Nieves-Rivera, Maria Ruizyantin & Michael D. Gottfried- 2003.
– The Miocene Climatic Optimum: evidence from ectothermic vertebratesof Central Europe – Palaeogeography, Palaeoclimatology, Palaeoecology195 (3-4): 389-401 – M. Böhme – 2003.
– Age of Carcharocles megalodon (Lamniformes:Otodontidae) : A reviewof the stratigraphic records – The Palaeontological Society of Japan (PSJ) (Japan) 75 (75): 7-15. – Hebe Hideo, Goto Mastatoshi &Kaneko Naotomo – 2004.
– Kæmpetandede hvide hajer og bredtandede makohajer (Lamnidae) fra det neogene område i Venezuela: Deres rolle i Caribien, Shallow-water FishAssemblage – Caribbean Journal of Science 40 (3): 362-368. – O.Aguilera & E. R. D. Aguilera – 2004.
– Tracing the ancestry of the Great White Shark – Journal of VertebratePaleontology 26 (4): 806-814 – K. G. Nyberg, C. N. Ciampaglio &G. A. Wray – 2006.
– Late Neogene Oceanographic Change along Florida’s West Coast:Evidence and Mechanisms – The Journal of Geology (USA: The Universityof Chicago) 104 (2): 143-162. – Warren D. Allmon, Steven D. Emslie,Douglas S. Jones & Gary S. Morgan – 2006.
– Tre-dimensionel computeranalyse af hvidhajens kæbemekanik: Hvor hårdt kan en hvidhaj bide? – Journal of Zoology 276 (4): 336-342. -S. Wroe, D. R. Huber, M. Lowry, C. McHenry, K. Moreno, P. Clausen, T.L. Ferrara, E. Cunningham, M. N. Dean & A. P. Summers – 2008.
– Miocæne hajer i Kendeace- og Grand Bay-formationerne i Carriacou,The Grenadines, Lesser Antilles – Caribbean Journal of Science. 44 (3)pp. 279-286. – Roger Portell, Gordon Hubell, Stephen Donovan, JeremyGreen, David Harper & Ron Pickerill – 2008.
– Giant-toothed white sharks and cetacean trophic interaction from thePliocene Caribbean Paraguaná Formation – Paläontologische Zeitschrift(Springer Berlin) 82 (2): 204-208. – Orangel A. Augilera, Luis García *Mario A. Cozzuol – 2008.
– Ancient Nursery Area for the Extinct Giant Shark Megalodonfrom theMiocene of Panama – PLoS ONE (Panama: PLoS.org) 5 (5): e10552 -Catalina Pimiento, Dana J. Ehret, Bruce J. Ehret, Bruce J. McFadden & GordonHubbell – 2010.
– The Great White Shark Carcharodon carcharias(Linne, 1758) in thePliocene of Portugal and its Early Distribution in Eastern Atlantic -Revista Española de Paleontología (Portugal) 25 (1): 1-6. – MiguelTelle Antunes, Ausenda Cáceres Balbino – 2010.
– Patterns and ecosystem consequences of shark declines in the ocean -Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055-1071. -Francesco Ferretti, Boris Worm, Gregory L. Britten, Michael J. Heithaus& Heike K. Lotze – 2010.
– Oprindelse af den hvide haj Carcharodon(Lamniformes: Lamnidae)baseret på en omkalibrering af den øvre neogene Pisco Formation fra Peru- Palaeontology 55(6):1139-1153 – D. J.Ehret, B.J. MacFadden, D. S. Jones, T. J. DeVries, D. A.Foster & R. Salas-Gismondi – 2012.
– Evolution of white and megatooth sharks, and evidence for earlypredation on seals, sirenians, and whales – Natural Science (CzechRepublic) 5 (11): 1203-1218. – C. G. Diedrich – 2013.
– Sharks and Rays (Chondrichthyes, Elasmobranchii) from the LateMiocene Gatun Formation of Panama – Journal of Paleontology 87 (5):755-774 – Catalina Pimiento, Gerardo González-Barba, Dana J. Ehret,Austin J. W. Hendy, Bruce J. MacFadden & Carlos Jaramillo -2013.
– Hvornår blev Carcharocles megalodon uddød? ANew Analysis of theFossil Record. – PLOS ONE. 9 (10): e111086. – C. Pimiento & C.F. Clements – 2014.
– Body-size trends of the extinct giant shark Carcharoclesmegalodon: adeep-time perspective on marine apex predators. – Paleobiology. 41 (3):479-490. – C. Pimiento & M. A. Balk – 2015.
– Optegnelser af Carcharocles megalodon i det østlige Guadalquivir-bassin (øvre Miocæn, Sydspanien). Estudios Geológicos. 71 (2): e032. – M.Reolid & J. M. Molina – 2015.
– Geografiske fordelingsmønstre af Carcharocles megalodonover tid afslører spor om uddøelsesmekanismer. – Journal ofBiogeography. 43 (8): 1645-1655. – C. Pimiento, B. J. MacFadden, C. F. Clements, S. Varela, C. Jaramillo, J. Velez-Juarbe & B. R. Silliman – 2016.
– The size of the megatooth shark, Otodus megalodon(Lamniformes:Otodontidae), revisited. – Historical Biology: 1-8. – Kenshu Shimada -2019.
– Den tidlige pliocæne uddøen af megatandhajen Otodusmegalodon: et synspunkt fra det østlige nordlige Stillehav. – PeerJ. 7:e6088. -R. W. Boessenecker, D. J. Ehret, D. J. Long, M. Churchill, E. Martin& S. J. Boessenecker – 2019.
– Overgangen mellem Carcharocles chubutensisog Carcharoclesmegalodon (Otodontidae, Chondrichthyes): lateral cusplet lossthroughtime. – Journal of Vertebrate Paleontology. 38 (6): e1546732. – V. J.Perez, S. J. Godfrey, B. W. Kent, R. E. Weems & J. R. Nance -2019.
– Body dimensions of the extinct giant shark Otodus megalodon:a 2Dreconconstruction. – Scientific Reports. 10 (14596): 14596. – J. A.Cooper, C. Pimiento, H. G. Ferrón & M. J. Benton – 2020.
—————————————————————————-
Uvilkårlige favoritter
 |
 |
 |
|
 |
Privatliv& Cookies-politik