Paranøddeindustrien — Fortid, nutid og fremtid
Scott A. Mori
The New York Botanical Garden
Tryk af denne artikel er sket med tilladelse fra Sustainable Harvest and Marketing of Rain Forest Products. Plotkin, M. og L. Famolare (eds.). Copyright Island Press, 1992. Udgivet af Island Press, Washington, D.C. & Covelo, Californien. For at bestille et eksemplar af denne bog, ring venligst 1-800-828-1302, eller skriv til: Island Press, Box 7, Covelo, CA 95428
Indledning | Naturhistorie | Fænologi | Bestøvningsbiologi | Spredningsbiologi | Paranøddehøst |
Brasanøddeplantationer | Fremtiden for paranøddeproduktion | Referencer
Indledning
Amazoniens skove rummer mange planter af økonomisk værdi (Balick, 1985; Farnsworth, 1984). Faktisk er intakte Amazonas-skove ofte mere værdifulde for deres træ- og andre produkter end de landbrugsplantager eller græsningsarealer, som alt for ofte erstatter dem (Peters et al., 1989; Menezes, 1990). Desuden har skovene i Amazonas en uvurderlig værdi som reservoirer for biodiversitet, regulatorer af de hydrologiske kredsløb, beskyttere af skrøbelige jorde og stabilisatorer af atmosfæren. På grund af værdien af skovene i Amazonas bør de, der ønsker at erstatte dem med landbrug eller græsningsarealer, være forpligtet til at påvise, at deres projekter vil give et udbytte, der er større end værdien af de intakte skove.
En af de vigtigste økonomiske planter i Amazonas er paranødden (Bertholletia excelsa, familie Lecythidaceae). De spiselige frø af denne art er sammen med latexen fra Hevea brasiliensis ofte nævnt som de vigtigste produkter fra udvindingsreserver i Amazonas. Paranødder indsamles hovedsagelig i den våde sæson, og gummi udvindes hovedsagelig i den tørre sæson. Kombinationen af disse to skovprodukter giver indkomst hele året rundt for dem, der lever af udvindingsindustrien. Indsamlingen af paranødder og gummi har en forholdsvis lille indvirkning på økologien i Amazonas’ skove. Derfor hævdes det ofte, at bevarelse af biodiversiteten og udnyttelse af disse og andre produkter, der ikke er træprodukter, er forenelige. De, der indsamler andre skovprodukter end træprodukter, er imidlertid næsten altid involveret i andre aktiviteter som f.eks. landbrug, træudvinding, minedrift og jagt. Som følge heraf beskytter stærkt udnyttede udvindingsreservater normalt kun en del af den enorme biodiversitet, der findes i Amazonas’ skove. Derfor skal enhver bevaringsplan for Amazonas omfatte store reserver af alle Amazonas-økosystemer, som er beskyttet mod overdreven økonomisk udnyttelse.
I dette kapitel gennemgår jeg den naturlige historie, værdien af høsten af paranødder, mulighederne for plantageopdyrkning og fremtiden for paranøddeindustrien. På grund af sin økonomiske betydning har paranødden været genstand for mange undersøgelser af dens biologi og agronomi. De fleste af disse undersøgelser er blevet udført i regi af “Centro de Pesquisa Agropecuária do Trópico Umido” (CPATU) under “Empresa Brasileira de Pesquisas Agropecuária” (EMBPRAPA) i Belém, Pará, Brasilien. Der findes en bibliografi om paranødder med 259 titler (Vaz Pereira og Lima Costa, 1981), og en nyere sammenfatning af paranøddernes biologi og agronomi findes i Mori og Prance (1990b).
Taksonomi og udbredelse
Bertholletia excelsa tilhører en pantropisk træfamilie (Lecythidaceae), der omfatter ca. 200 arter i Neotropien, fordelt fra det sydlige Mexico til det sydlige Brasilien (Mori og Prance, 1990a; Prance og Mori, 1979). Paranødden udgør en enkelt art i den veldefinerede slægt Bertholletia. Selv om der er betydelig variation i frugtstørrelse og -form og antallet af frø pr. frugt, er der ingen begrundelse for at anerkende mere end én art af Bertholletia.
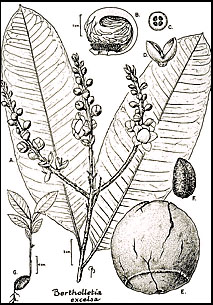 De nærmeste slægtninge til B. excelsa er blandt arter af Lecythis, der i folkemunde almindeligvis omtales som jarana-gruppen (Mori og Prance, 1990b). Andre arter med spiselige frø i familien er Lecythis pisonis og dens slægtninge, L. minor og L. ollaria. De to sidstnævnte arter forårsager dog undertiden hår- og negletab på grund af overskydende selenophobning i frøene (Dickson, 1969; Kerdel-Vegas, 1966). Ikke desto mindre er forskellene mellem Bertholletia og Lecythis så store, at der er ringe håb om at indføre genetisk materiale fra den ene slægt til den anden via hybridisering. Derfor må kimplasma til forbedring af paranøddeproduktionen komme fra den variation, der findes inden for B. excelsa, og ikke fra nært beslægtede arter i andre slægter.
De nærmeste slægtninge til B. excelsa er blandt arter af Lecythis, der i folkemunde almindeligvis omtales som jarana-gruppen (Mori og Prance, 1990b). Andre arter med spiselige frø i familien er Lecythis pisonis og dens slægtninge, L. minor og L. ollaria. De to sidstnævnte arter forårsager dog undertiden hår- og negletab på grund af overskydende selenophobning i frøene (Dickson, 1969; Kerdel-Vegas, 1966). Ikke desto mindre er forskellene mellem Bertholletia og Lecythis så store, at der er ringe håb om at indføre genetisk materiale fra den ene slægt til den anden via hybridisering. Derfor må kimplasma til forbedring af paranøddeproduktionen komme fra den variation, der findes inden for B. excelsa, og ikke fra nært beslægtede arter i andre slægter.
Bertholletia excelsa er en plante fra Amazonas, som foretrækker ikke oversvømmede skove (terra firma) i Guianas, Colombia, Venezuela, Peru, Bolivia og Brasilien. De klimatiske forhold, under hvilke paranødder vokser, er opsummeret i de Almeida (1963), Diniz og Bastos (1974) og Mori og Prance (1990b).
Brasilienødder dyrkes i tropiske botaniske haver langt uden for dens oprindelige udbredelsesområde, og der er etableret mindre plantager i Kuala Lumpur i Malaysia (Müller, 1981) og Ghana i Afrika (D.K. Abbiw, pers. comm.).
Træer af Bertholletia excelsa forekommer i bestande på 50 til 100 individer, der er kendt som “manchales” i Peru (Sánchez, 1973) og “castanhais” i Brasilien (Dias, 1959). Tætheden af paranøddetræer pr. hektar varierer betydeligt i hele Amazonasområdet. I en undersøgelse af produktionen af paranødder i det østlige Brasilien fandt Miller (1990) mellem 9 og 26 reproduktive træer pr. hektar, mens Becker og Mori (upublicerede data) kun fandt ét træ på over 10 cm dbh i et 100 hektar stort område i det centrale Amazonas Brasilien.
Der er visse tegn på, at paranøddetræer er afstandsafhængige (Mori og Prance, 1990b). Desuden har nogle forfattere foreslået, at bevoksninger af paranøddetræer har deres oprindelse hos de prækolombianske indianere (Miller, 1990; Mori og Prance, 1990b; Müller et al., 1980). Der er stadig behov for en forståelse af udviklingen af reproduktive individer fra frø, før det er muligt at forvalte paranødder i naturlige bestande.
Blomning af Bertholletia excelsa finder sted i løbet af den tørre sæson og ind i den våde sæson. Paranødder vokser faktisk kun naturligt i regioner med en tre til fem måneder lang tørtid (Müller, 1981). I den østlige del af det brasilianske Amazonasområde begynder blomstringen ved slutningen af regntiden i september og strækker sig til februar. Den maksimale blomstring finder sted i oktober, november og december (Moritz, 1984).
Ved slutningen af regntiden, som regel i juli, begynder bladene på paranøddetræer at falde. Den nye vækst skyller ud lige under det foregående års blomsterstande, og de nye blomsterstande produceres i toppen af den aktuelle vækstflush. Der produceres et stort antal blomster dagligt over en forholdsvis lang periode. Blomsterne åbner sig mellem kl. 4.30 og 5.00 om morgenen. Blomsternes støvknapper begynder dog at afblødes i knoppen flere timer før blomsterne åbner sig. Kronbladene og androecia falder om eftermiddagen på den dag, hvor blomsterne åbner sig (Mori og Prance, 1990b).
Frugtudviklingen tager længere tid hos B. excelsa end hos nogen anden art af Lecythidaceae. Moritz (1984) angiver, at det tager 15 måneder for frugterne at udvikle sig, efter at de er blevet sat. Følgelig falder paranøddernes frugter for det meste i januar og februar i løbet af regntiden. Under naturlige forhold tager det 12 til 18 måneder for frøene at spire (Müller, 1981).
 Paranøddens blomster er zygomorfe med et androecium, der på den ene side er forlænget til en hætte, der buer sig over og er tæt knyttet til toppen af æggestokkene. Desuden sidder kronbladene fast på androeciumet (Fig. 27-1). Derfor kan blomsterne kun komme ind i blomsterne af store bier med tilstrækkelig styrke til at bryde androecialhætten op for at få den bestøverbelønning, som formodes at være nektar, der produceres i spidsen af den sammenrullede androecialhætte. Bier af slægterne Bombus, Centris, Epicharis, Eulaema og Xylocopa er blevet fanget på besøg hos paranøddetræer (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985). Disse bier er ikke-sociale eller halvsociale og kan derfor ikke let manipuleres af mennesker, som det er tilfældet med de sociale bier (f.eks. Apis, Melipona og Trigona), der kan bruges til at bestøve visse afgrøder ved at transportere bistader fra den ene plantage til den anden.
Paranøddens blomster er zygomorfe med et androecium, der på den ene side er forlænget til en hætte, der buer sig over og er tæt knyttet til toppen af æggestokkene. Desuden sidder kronbladene fast på androeciumet (Fig. 27-1). Derfor kan blomsterne kun komme ind i blomsterne af store bier med tilstrækkelig styrke til at bryde androecialhætten op for at få den bestøverbelønning, som formodes at være nektar, der produceres i spidsen af den sammenrullede androecialhætte. Bier af slægterne Bombus, Centris, Epicharis, Eulaema og Xylocopa er blevet fanget på besøg hos paranøddetræer (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985). Disse bier er ikke-sociale eller halvsociale og kan derfor ikke let manipuleres af mennesker, som det er tilfældet med de sociale bier (f.eks. Apis, Melipona og Trigona), der kan bruges til at bestøve visse afgrøder ved at transportere bistader fra den ene plantage til den anden.
 For det meste er krydsbestøvning nødvendig for frøsætningen hos neotropiske Lecythidaceae. Derfor er bierne og i mindre grad flagermusene afgørende for bestøvningen og den efterfølgende udvikling af frugter og frø hos Lecythidaceae. Selv om der kan forekomme et lavt niveau af indavl hos Bertholletia excelsa, er det meste af frøsætningen hos denne art et resultat af krydsbestøvning (Mori og Prance, 1990b). Udviklingen af selvkompatible linjer af paranødden ville lette plantagedyrkningen af denne art ved at eliminere behovet for krydsbestøvning fra de vanskeligt håndterbare bi-pollinatorer.
For det meste er krydsbestøvning nødvendig for frøsætningen hos neotropiske Lecythidaceae. Derfor er bierne og i mindre grad flagermusene afgørende for bestøvningen og den efterfølgende udvikling af frugter og frø hos Lecythidaceae. Selv om der kan forekomme et lavt niveau af indavl hos Bertholletia excelsa, er det meste af frøsætningen hos denne art et resultat af krydsbestøvning (Mori og Prance, 1990b). Udviklingen af selvkompatible linjer af paranødden ville lette plantagedyrkningen af denne art ved at eliminere behovet for krydsbestøvning fra de vanskeligt håndterbare bi-pollinatorer.
Bier uden for paranøddens oprindelige udbredelsesområde kan foretage bestøvning. F.eks. sætter paranøddetræer på Ceylon (Macmillan, 1935), i Kuala Lumpur og Ghana frugt. Det vides imidlertid ikke, om bestøvere uden for paranøddens oprindelige udbredelsesområde eller “ukrudtsbestøvere”, der findes i sekundære skove, er effektive nok til at muliggøre en økonomisk levedygtig frugtproduktion.
 Frugterne og frøene af B. excelsa er unikke i Lecythidaceae. Ved modenhed falder de store, runde, træagtige frugter ned på jorden med frøene indeni. Frøene, som har en knoglet testa, fjernes fra kapslerne og spredes af gnavere, især agoutis (Dasyprocta spp.) (Huber, 1910; Miller; 1990; E. Ortiz, pers. comm.). Agoutis og egern er muligvis de eneste dyr, der er i stand til effektivt at gnave sig gennem de ekstremt træagtige pericarps. De spiser nogle af frøene og gemmer andre til senere brug. Følgelig bliver frøene enten spist og ødelagt, eller de bliver efterladt i et glemt gemmested, hvor de til sidst kan spire 12 til 18 måneder senere.
Frugterne og frøene af B. excelsa er unikke i Lecythidaceae. Ved modenhed falder de store, runde, træagtige frugter ned på jorden med frøene indeni. Frøene, som har en knoglet testa, fjernes fra kapslerne og spredes af gnavere, især agoutis (Dasyprocta spp.) (Huber, 1910; Miller; 1990; E. Ortiz, pers. comm.). Agoutis og egern er muligvis de eneste dyr, der er i stand til effektivt at gnave sig gennem de ekstremt træagtige pericarps. De spiser nogle af frøene og gemmer andre til senere brug. Følgelig bliver frøene enten spist og ødelagt, eller de bliver efterladt i et glemt gemmested, hvor de til sidst kan spire 12 til 18 måneder senere.
 Paranødder høstes næsten udelukkende fra vilde træer i løbet af en periode på fem til seks måneder i regntiden. Frugterne, som vejer fra 0,5 til 2,5 kg og indeholder 10 til 25 frø, indsamles umiddelbart efter, at de er faldet ned, for at minimere insekt- og svampeangreb på frøene og for at kontrollere antallet af frø, der bæres væk af dyr (Mori og Prance, 1990b). Ifølge Miller (1990) varierer antallet af kapsler, der produceres pr. træ, fra 63 til 216. Mere detaljerede beskrivelser af metoderne til høst af paranødder kan findes i Almeida (1963), Mori og Prance (1990b) og Souza (1963).
Paranødder høstes næsten udelukkende fra vilde træer i løbet af en periode på fem til seks måneder i regntiden. Frugterne, som vejer fra 0,5 til 2,5 kg og indeholder 10 til 25 frø, indsamles umiddelbart efter, at de er faldet ned, for at minimere insekt- og svampeangreb på frøene og for at kontrollere antallet af frø, der bæres væk af dyr (Mori og Prance, 1990b). Ifølge Miller (1990) varierer antallet af kapsler, der produceres pr. træ, fra 63 til 216. Mere detaljerede beskrivelser af metoderne til høst af paranødder kan findes i Almeida (1963), Mori og Prance (1990b) og Souza (1963).
Indsamling af paranødder har stor betydning for de lokale økonomier i Amazonas. De tilgængelige tal giver dog kun en tilnærmelse af den samlede produktion, fordi det er vanskeligt at få nøjagtige data fra Amazonasområdet. Den brasilianske produktion har varieret fra 3 557 tons i 1944 til 104 487 tons i 1970. Siden 1980 har den årlige produktion ligget på omkring 40.000 tons (Mori og Prance, 1990b). Tidligere var velfærden i mange byer i Amazonas som Puerto Maldonado i Peru (Sánchez, 1973) og Marabá i Brasilien (Dias, 1959) i høj grad afhængig af produktionen af paranødder. I 1986 var den samlede værdi af afskallede og ikke-afskallede paranøddefrø, der blev eksporteret fra Manaus alene, 5 773 228 USD. (Mori og Prance, 1990b). De fleste af frøene sendes til England, Frankrig, USA og Tyskland.
Beregninger foretaget af Miller (1990) har anslået den primære værdi (penge betalt til samlerne) af paranøddebevoksninger til 97 $ pr. ha. Denne værdi omfatter en vilkårlig rabat på 25 % for at tage højde for frø, der er tilbage i bevoksningerne. Den sekundære værdi — med andre ord de penge, som det eksporterende selskab modtager for uskallede nødder fra et amerikansk importfirma — blev anslået til 175,56 $ pr. ha. Den tertiære værdi – prisen for en pose uskrællede nødder i et supermarked i Florida – blev beregnet til 1059,44 USD pr. ha. Skallede og forarbejdede nødder er meget mere værdifulde. Bestræbelser, som f.eks. fra Cultural Survival, for at placere noget af afskallingen og noget af forarbejdningen i samlernes lokalsamfund giver yderligere incitamenter til at bevare udvindingsreserver. I løbet af en tiårig periode synes udnyttelsen af en skov til produktion af paranødder at være mere rentabel end udvinding af tømmer eller hugst af skoven til græsningsarealer (Miller, 1990).
Paranøddeplantager
Metoden til dyrkning af paranødder i store plantager er blevet udviklet af Müller og hans medarbejdere fra CPATU, Belém, Brasilien. Et engelsk resumé af deres arbejde findes i Mori og Prance (1990b). Som tidligere nævnt indsamles det meste af produktionen af paranødder stadig fra vilde træer. Der er imidlertid ved at blive udviklet plantager i forskellige dele af Amazonas. I januar 1990 besøgte jeg Fazenda Aruanã, en paranøddeplantage, der ligger ved kilometer 215 på Manaus/Itacoatiara-vejen i staten Amazonas, Brasilien. Der er tale om en 12.000 hektar stor tidligere kvægfarm, der delvis blev omdannet til paranøddeplantage i 1980. På tidspunktet for mit besøg var der plantet 318.660 paranøddetræer på 3.341 hektar. Derudover var der et ukendt antal individer, der voksede i de nærliggende skove, som tilhørte Fazenda Aruanã.
Den oprindelige hensigt på Fazenda Aruanã var at plante paranøddetræer i gitre på 20 gange 20 meter og lade kvæg græsse mellem træerne. I begyndelsen spiste kvæget ikke bladene fra træerne, men senere gjorde de det. Som følge heraf blev afstanden reduceret til 10 gange 10 meter, og antallet af kvæg blev reduceret til 300 stykker. Træerne i Aruanã-plantagen er resultatet af podning af højtydende kloner fra Abufari-området i Amazonas, hvor paranødder er kendt for deres store frugter og frø. En vigtig overvejelse i forbindelse med etableringen af denne og andre plantager er frøenes oprindelse. Moritz (1984) har vist, at frugtproduktionen som følge af befrugtning mellem træer af samme klon er lav. På grundlag af dette koncept anbefaler Müller (1981), at knopper til podning bør hentes fra mindst fem forskellige træer. En fare ved at anvende så få kloner til podning er, at en udvælgelse med henblik på et højt frugtudbytte også kan reducere plantagens evne til at modstå fremtidige angreb af sygdomme og insekter.
Rodstokke dyrkes i Fazenda Aruanã fra frø. Frøene spirer ved at blødgøre dem i fugtigt sand og derefter fjerne embryoet ved at åbne frøskallen langs kanterne. Embryoerne, som er behandlet med et fungicid, spirer på ca. 20 dage, og frøplanterne dyrkes i plastikposer eller kopper. Når frøplanterne plantes ud på marken, tilsættes der ca. 200 g fosfor til det hul, hvori de plantes. Rodstokke er klar til podning efter ca. 1,5 til 2 år, når de unge træer er blevet veletableret i marken. Man opnår bedre podestammer, hvis bladene fjernes fra grenene 8 til 10 dage før podestammen fjernes. Dette fremmer dannelsen af kraftige knopper, som bedre tåler podning. Tredive dage efter, at podningen er foretaget, ringmærkes rodstokken distalt i forhold til podningen. Dette gør det muligt for podestammen at vokse gradvist ud af grenene på grundstammen. Efter seks år begynder træerne at bære frugt. På tidspunktet for mit besøg var alle de frugter, der indtil da var blevet produceret, dog blevet brugt til at dyrke mere rodstokke eller til at producere frøplanter til salg til andre.
Plantagen ser ikke ud til at have problemer som følge af manglen på bestøvere. Bixa orellana var blevet plantet for at levere pollen som fødekilde for bestøvere. Desuden er der udstrakt naturskov omkring plantagen, og vindmøller med naturlig vegetation gennemskæres plantagen. I disse møller findes der mange planter, som er kendt som fødekilder for bestøvere af paranøddetræer. Selv om træerne var næsten 10 år gamle på det tidspunkt, var der ingen tegn på eller historik med sygdom.
Ejerne af Fazenda Aruanã har store forventninger til denne plantage. Ved udgangen af 12 år forventer de et udbytte på 4,8 kg pr. træ og 480 dollars pr. ha. Efter 18 år forventer de et udbytte på 8,5 kg pr. træ og 850 USD pr. ha.
Fremtiden for produktionen af paranødder
Den hidtidige produktion af paranødder er hovedsagelig blevet indsamlet fra vilde træer. I de seneste år er produktionen af paranødder faldet på grund af skovrydning, paranøddesamleres udvandring til store metropoler, oversvømmelse af nogle traditionelle paranøddebevoksninger og måske på grund af forstyrrelser af bestøvere forårsaget af brande i den tørre sæson, hvor paranøddetræerne er i blomst (Kitamura og Müller, 1984; Mori og Prance, 1990b). Hvis de traditionelle metoder til produktion af paranødder skal opretholdes, er det nødvendigt at oprette meget store udvindingsreserver i områder med høj paranøddetræstæthed.
Den, der er interesseret i bevarelse af den tropiske natur, bør imidlertid være forsigtig med ikke at sætte lighedstegn mellem oprettelse af udvindingsreservater og bevarelse af Amazonas’ biodiversitet. Fordi samlere af paranødder og gummiklippere gør mere end blot at samle paranødder, har de ofte en negativ indvirkning på plante- og dyrediversiteten. Faktisk kan udvindingsreservater blive til lidt mere end sekundær vegetation med økonomiske planter som paranødder og gummitræer spredt her og der. Derfor ophæver oprettelsen af udvindingsreservater ikke behovet for velplanlagte biologiske reservater.
Den fremtidige succes for paranøddeplantager er stadig åben for diskussion. Indtil nu har der ikke været eksempler på økonomisk vellykkede plantager af dette træ fra Amazonas. Alle forsøg på at dyrke tropiske træer i plantager, som ikke naturligt vokser i næsten monotypiske bestande, har været en fiasko. I deres naturlige levested er paranøddetræer fordelt mere eller mindre spredt i skoven på samme måde som gummitræer. Den økonomiske katastrofe ved at forsøge at indføre gummi i plantager inden for dens hjemområde er blevet veldokumenteret (Hecht og Cockburn, 1989), og der er ingen grund til at tro, at paranøddeplantager i Amazonas ikke vil gå samme skæbne i møde som de amazoniske gummiplantager. En omhyggelig observation af Aruanã-plantagen i løbet af det næste årti kan give os mulighed for at afgøre, om produktion af paranødder i plantager er et levedygtigt alternativ til indsamling fra vilde træer. Hvis plantagerne er levedygtige, må naturbevarere være forberedt på at vurdere den indvirkning, som plantagerne vil have på opretholdelsen af udvindingsreserver.
Endeligt er det vigtigt, at vi ikke sætter for store forhåbninger til udvinding af paranødder som en økonomisk levedygtig måde at forsørge en stadig stigende befolkning i Amazonas på. For det første vil verdensmarkederne måske ikke være i stand til at håndtere en stor stigning i produktionen af paranødder, og for det andet er en så lav intensitetsudnyttelse af jorden ikke i stand til at understøtte den menneskelige befolkning på det niveau, der er nødvendigt for at øge den levestandard, som flere og flere mennesker efterspørger. Hvis Ewels (1991) estimater om, at jagt-samler- og skiftende landbrug kun kan forsørge henholdsvis én person pr. 5 kvadratkilometer og én person pr. kvadratkilometer, er korrekte (der er ingen grund til at tro, at de ikke er det), så vil udvindingsreserverne kun gøre meget lidt for at absorbere befolkningstilvæksten i Amazonaslandene. Fremtiden for udvindingsreservater og menneskehedens evne til at bevare et repræsentativt udsnit af den neotropiske biodiversitet afhænger af, om det lykkes at kontrollere befolkningstilvæksten og forbruget både inden for og uden for troperne (Erhlich og Erhlich, 1990).
de Almeida, C.P. 1963. Castanha do Para: Sua exportação e importância na economia Amazonica. Edições S.I.A. Estudos Brasileiros 19:1-86.
Balick, M.J. 1985. Useful Plants of Amazonia: A Resource of Global Importance. I G.T. Prance og T.E. Lovejoy (eds.), Amazonia. New York: Pergamon Press.
Dias, C.V. 1959. Aspectos geográficos do comércio da castanha no médio Tocantins. Revista Brasil. Geogr. 21(4):77-91.
Dickson, J.D. 1969. Bemærkninger om hår- og negletab efter indtagelse af Sapucaia-nødder (Lecythis elliptica). Econ. Bot. 23:133-34.
de Diniz, T.D., A. S. Diniz, og T. X. Bastos. 1974. Contribuição ao conhecimento do clima tipico da castanha do Brasil. Bol. Técn. IPEAN 64:59-71.
Ewel, J. 1991. Bevaring og landbrug. Tropinet 2(1):1.
Ehrlich, A.H., og P.R. Ehrlich. 1990. Udryddelse: Life in Peril. I S. Head og R. Heinzman (eds.), Lessons of the Rainforest. San Francisco: Sierra Club Books.
Farnsworth, N.R. 1984. Hvordan kan brønden være tør, når den er fyldt med vand? Econ. Bot. 38:4-13.
Hecht, S., og A. Cockburn. 1989. The Fate of the Forest. New York: Verso.
Huber, J. 1910. Mattas e madeiras amazônicas. Bol. Mus. Paraense Hist. Nat. 6:91-225.
Kerdel-Vegas, F. 1966. Den hårfjerningsvirkende og cytotoksiske virkning af Coco De Mono (Lecythis ollaria) og dens relation til kronisk seleniose. Econ. Bot. 20:187-95.
Kitamura, P.C., og C.H. Müller. 1984. Castanhais nativas de Marabá-PA: Fatores de depredação e bases para a sua preservação. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Documentos 30:1-32.
Macmillan, H.F. 1935. Tropical Planting and Gardening with Special Reference to Ceylon. London: Macmillan.
Menezes, M.A. 1990. (Reservas extrativistas: Por uma reforma agrária ecológica.) Ciência Hoje 11(64):4-6.
Miller, C. 1990. Natural History, Economic Botany, and Germplasm Conservation of the Brazil Nut Tree (Bertholletia excelsa Humb. and Bonpl). Masterafhandling fremlagt ved University of Florida.
Mori, S. A., og G.T. Prance. 1990a. Lecythidaceae–Part II. The Zygomorphic-Flowered New World Genera (Bertholletia, Corythophora, Couratari, Couroupita, Eschweilera, and Lecythis). Fl. Neotrop. Monogr. 21(II): 1-376.
Mori, S. A., og G.T. Prance. 1990b. Taxonomy, Ecology, and Economic Botany of the Brazil nut (Bertholletia excelsa Humb. and Bonpl.: Lecythidaceae). Adv. Econ. Bot. 8:130-50.
Moritz, A. 1984. Biologiske undersøgelser af paranødden (Bertholletia excelsa H.B.K.). EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Dokumenter 29:1-82.
Müller, C. H. 1981. paranødder; agronomiske undersøgelser. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Dokumenter 2: 1-25.
Müller, C. H., I. A. Rodriques, A. A. Müller, og N.R.M. Müller. 1980. Paranødder. Forskningsresultater. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Diverse 2:1-25.
Nelson, B. W., M. L. Absy, E. M. Barbosa, og G. T. Prance. 1985. Iagttagelser om blomsterbesøgende på Bertholletia excelsa H.B.K. og Couratari tenuicarpa A. C. Sm. (Lecythidaceae). Acta Amazonia 15 (1/2):225-34.
Peters, C.M., A. H. Gentry, og R.O. Mendelsohn. 1989. Værdiansættelse af en amazonisk regnskov. Nature 339:655-56.
Prance, G.T., og S.A. Mori. 1979. Lecythidaceae–Del I. De aktinomorfe blomstrede Lecythidaceae fra den nye verden (Asteranthos, Gustavia, Grias, Allantoma og Cariniana). Fl. Neotrop. Monogr. 21(1):1-270.
Sánchez. 1973. Udnyttelse og afsætning af paranødder i Madre de Dios. Landbrugsministeriet, Generaldirektoratet for Skovbrug og Jagt, rapport nr. 20. Lima, Peru.
Souza A. H. 1963. Castanha do Pará: Estudo botánico químico e tecnológico. Ediçoes S.I.A., Estudos Técnicos 23:1-69.
Vaz Pereira, I.C., og S.L. Lima Costa. 1981. Bibliografia de Castanha-do-Pará (Bertholletia excelsa H.B.K.). EMPRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Belém, Pará.
Vend tilbage til Lecythidaceae Pages Home
Vend tilbage til Mori Home
Etableret februar 2002. Hvis du har problemer eller spørgsmål, bedes du kontakte [email protected].