Evolution und Natürliche Auslese
Die Natur ermutigt keine Lockerheit, sie verzeiht keine Fehler
– Ralph Waldo Emerson
Ich habe dieses Prinzip, durch das jede geringfügige Veränderung, wenn sie nützlich ist, erhalten wird, mit dem Begriff Natürliche Auslese bezeichnet.
– Charles Darwin, Die Entstehung der Arten
|
10. Oktober 2010 |
|
In dieser Lektion wollen wir fragen:
- Wie haben Beobachtungen in der Natur zur Formulierung der Evolutionstheorie geführt?
- Was sind die wichtigsten Punkte von Darwins Evolutionstheorie?
- Wie funktioniert der Prozess der natürlichen Selektion?
- Welche Beweise gibt es für lokale Anpassung?
- Wie kann die natürliche Auslese die Häufigkeit von Merkmalen über aufeinanderfolgende Generationen hinweg beeinflussen?
Die (R)Evolution der Theorie
Die Evolutionstheorie ist eine der großen intellektuellen Revolutionen der Menschheitsgeschichte, die unsere Wahrnehmung der Welt und unseres Platzes in ihr drastisch verändert hat. Charles Darwin stellte eine kohärente Evolutionstheorie auf und sammelte eine Fülle von Beweisen für diese Theorie. Zu Darwins Zeiten glaubten die meisten Wissenschaftler fest daran, dass jeder Organismus und jede Anpassung das Werk eines Schöpfers war. Linneaus führte das System der biologischen Klassifizierung ein, das wir heute verwenden, und er tat dies im Geiste der Katalogisierung von Gottes Schöpfungen.
Mit anderen Worten: Alle Ähnlichkeiten und Unterschiede zwischen den Organismengruppen, die das Ergebnis des Verzweigungsprozesses sind, der den großen Baum des Lebens hervorbringt (siehe Abbildung 1), wurden von den Philosophen und Wissenschaftlern des frühen 19. Jahrhunderts als Folge eines allmächtigen Entwurfs angesehen.
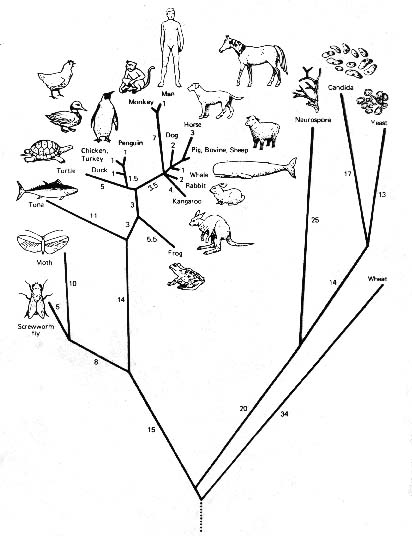
Abbildung 1: Ein phylogenetischer „Lebensbaum“, der durch Computeranalyse der Cyochrom-c-Moleküle in den abgebildeten Organismen erstellt wurde; es gibt so viele verschiedene Lebensbäume wie es Analysemethoden zu ihrer Erstellung gibt.
Im 19. Jahrhundert begannen jedoch einige Naturhistoriker, den evolutionären Wandel als Erklärung für die in der Natur beobachteten Muster in Betracht zu ziehen. Die folgenden Ideen waren Teil des intellektuellen Klimas zur Zeit Darwins.
- Niemand wusste, wie alt die Erde war, aber Geologen begannen zu schätzen, dass die Erde wesentlich älter war, als es die biblische Schöpfung erklärte. Die Geologen lernten mehr über Schichten, die durch die Ablagerung von Sedimenten in aufeinander folgenden Perioden entstanden. Dies legte eine zeitliche Abfolge nahe, bei der jüngere Schichten über älteren Schichten lagen.
- Das Konzept des Uniformitarismus, das vor allem auf den einflussreichen Geologen Charles Lyell zurückgeht, versuchte, die Erdgeschichte zu entschlüsseln, und zwar unter der Arbeitshypothese, dass die gegenwärtigen Bedingungen und Prozesse der Schlüssel zur Vergangenheit sind, indem laufende, beobachtbare Prozesse wie Erosion und die Ablagerung von Sedimenten untersucht wurden.
- Im 18. und 19. Jahrhundert häuften sich die Entdeckungen von Fossilien. Zunächst dachten die Naturforscher, sie fänden die Überreste unbekannter, aber noch lebender Arten. Als die Fossilienfunde jedoch weitergingen, wurde deutlich, dass von keinem Ort der Erde etwas Ähnliches wie riesige Dinosaurier bekannt war. Außerdem wies Cuvier bereits um 1800 darauf hin, dass die Fossilien den existierenden Arten umso weniger ähnelten, je tiefer die Schichten lagen.
- Ähnlichkeiten zwischen Organismengruppen galten als Beweis für Verwandtschaft, was wiederum auf evolutionäre Veränderungen schließen ließ. Darwins intellektuelle Vorgänger akzeptierten die Idee evolutionärer Beziehungen zwischen Organismen, konnten aber keine zufriedenstellende Erklärung dafür liefern, wie die Evolution stattfand.
- Lamarck ist der berühmteste von ihnen. Im Jahr 1801 schlug er die organische Evolution als Erklärung für die physische Ähnlichkeit zwischen Gruppen von Organismen vor und schlug einen Mechanismus für adaptive Veränderungen vor, der auf der Vererbung erworbener Merkmale beruht. Er schrieb über die Giraffe:
„Wir wissen, dass dieses Tier, das größte aller Säugetiere, im Innern Afrikas lebt, an Orten, wo der Boden, der fast immer trocken und ohne Gras ist, sie zwingt, auf Bäumen zu grasen und sich ständig anzustrengen, um sie zu erreichen. Diese lang anhaltende Gewohnheit hat bei allen Mitgliedern ihrer Rasse dazu geführt, dass die Vorderbeine länger geworden sind als die Hinterbeine und dass ihr Hals so gestreckt ist, dass die Giraffe, ohne sich auf die Hinterbeine zu stellen, ihren Kopf bis zu einer Höhe von sechs Metern hebt.“
Dies besagt im Wesentlichen, dass die Hälse der Giraffen durch die ständige Streckung, um hohes Laub zu erreichen, lang wurden. Larmarck lag mit dem angenommenen Mechanismus natürlich falsch, aber sein Beispiel macht deutlich, dass Naturforscher in den frühen 1800er Jahren über die Möglichkeit evolutionärer Veränderungen nachdachten.
- Darwin wurde durch Beobachtungen beeinflusst, die er während seiner Jugendreise als Naturforscher auf dem Vermessungsschiff Beagle machte. Auf den Galapagos-Inseln bemerkte er die leichten Unterschiede, die Schildkröten von verschiedenen Inseln erkennbar machten. Er beobachtete auch eine ganze Reihe einzigartiger Finken, die berühmten „Darwinfinken“, die sich von Insel zu Insel leicht unterschieden. Außerdem schienen sie alle dem gemeinen Finken auf dem Festland von Ecuador, 600 Meilen östlich, zu ähneln, sich aber von ihm zu unterscheiden. Muster in der Verteilung und Ähnlichkeit von Organismen hatten einen wichtigen Einfluss auf Darwins Denken. Das Bild oben auf dieser Seite ist von Darwins eigenen Skizzen von Finken in seinem Journal of Researches.
- 1859 veröffentlichte Darwin sein berühmtes Werk Über die Entstehung der Arten durch natürliche Auslese, einen Wälzer von über 500 Seiten, in dem er umfangreiche Beweise für seine Theorie zusammenstellte. Die Veröffentlichung des Buches löste einen Sturm der Entrüstung aus – jedes Exemplar des Buches wurde am Tag seiner Veröffentlichung verkauft. Mitglieder der religiösen Gemeinschaft sowie einige wissenschaftliche Kollegen waren über Darwins Ideen empört und protestierten. Die meisten Wissenschaftler erkannten jedoch die Kraft von Darwins Argumenten. Noch heute streiten sich die Schulbehörden über die Gültigkeit und die Eignung von Darwins Theorie für die Lehrpläne der Naturwissenschaften, und um die Kontroverse herum hat sich eine ganze Reihe von Debatten entwickelt (siehe die WWW-Site Talk.Origins für einen laufenden Dialog). Wir haben nicht die Zeit, um alle Beweise und Argumente Darwins zu behandeln, aber wir können die Kernideen untersuchen. Was besagt diese Evolutionstheorie?
Darwins Theorie
Darwins Evolutionstheorie beinhaltet die folgenden Grundgedanken. Die ersten drei Ideen wurden bereits von früheren und gleichzeitigen Naturforschern diskutiert, die sich mit dem „Artenproblem“ beschäftigten, als Darwin seine Forschungen begann. Darwins ursprüngliche Beiträge waren der Mechanismus der natürlichen Selektion und eine Fülle von Beweisen für evolutionäre Veränderungen aus vielen Quellen. Er lieferte auch durchdachte Erklärungen zu den Folgen der Evolution für unser Verständnis der Geschichte des Lebens und der modernen biologischen Vielfalt.
-
Spezies (Populationen sich kreuzender Organismen) verändern sich im Laufe der Zeit und des Raums. Die Vertreter der heute lebenden Arten unterscheiden sich von denen, die in der jüngeren Vergangenheit lebten, und Populationen in verschiedenen geografischen Regionen unterscheiden sich heute leicht in Form oder Verhalten. Diese Unterschiede reichen bis in den Fossilienbestand hinein, der diese Behauptung reichlich untermauert.
-
Alle Organismen haben gemeinsame Vorfahren mit anderen Organismen. Im Laufe der Zeit können sich Populationen in verschiedene Arten aufspalten, die eine gemeinsame Vorfahrenpopulation haben. Wenn man weit genug in der Zeit zurückgeht, hat jedes Organismenpaar einen gemeinsamen Vorfahren. So hatten beispielsweise Menschen und Schimpansen vor etwa acht Millionen Jahren, Wale vor etwa 60 Millionen Jahren und Kängurus vor über 100 Millionen Jahren einen gemeinsamen Vorfahren. Die gemeinsame Abstammung erklärt die Ähnlichkeiten von Organismen, die zusammen klassifiziert werden: ihre Ähnlichkeiten spiegeln die Vererbung von Merkmalen von einem gemeinsamen Vorfahren wider.
-
Evolutionäre Veränderungen sind nach Darwins Ansicht allmählich und langsam. Diese Behauptung wurde durch die langen Episoden allmählicher Veränderungen bei Organismen im Fossilbericht und die Tatsache gestützt, dass zu Darwins Zeiten kein Naturforscher das plötzliche Auftreten einer neuen Art beobachtet hatte. Seitdem haben Biologen und Paläontologen ein breites Spektrum von langsamen bis schnellen evolutionären Veränderungen innerhalb von Tierlinien dokumentiert.
Der wichtigste Mechanismus für Veränderungen im Laufe der Zeit ist die natürliche Auslese, die im Folgenden näher erläutert wird. Dieser Mechanismus bewirkt, dass sich die Eigenschaften (Merkmale) von Organismen innerhalb von Abstammungslinien von Generation zu Generation verändern.
Der Prozess der natürlichen Selektion
Darwins Prozess der natürlichen Selektion hat vier Komponenten.
- Variation. Organismen (innerhalb von Populationen) weisen individuelle Variationen in Aussehen und Verhalten auf. Diese Variationen können die Körpergröße, die Haarfarbe, die Gesichtszeichnung, die Stimme oder die Anzahl der Nachkommen betreffen. Andererseits gibt es Merkmale, die zwischen Individuen wenig bis gar nicht variieren, wie zum Beispiel die Anzahl der Augen bei Wirbeltieren.
- Vererbung. Einige Merkmale werden konsequent von den Eltern an die Nachkommen weitergegeben. Solche Merkmale sind vererbbar, während andere Merkmale stark von Umweltbedingungen beeinflusst werden und eine schwache Vererbbarkeit aufweisen.
- Hohe Wachstumsrate der Population. Die meisten Populationen haben jedes Jahr mehr Nachkommen als die lokalen Ressourcen tragen können, was zu einem Kampf um Ressourcen führt. Jede Generation erfährt eine beträchtliche Sterblichkeit.
- Differenzielles Überleben und Reproduktion. Individuen mit Merkmalen, die für den Kampf um lokale Ressourcen gut geeignet sind, werden mehr Nachkommen zur nächsten Generation beitragen.
Von einer Generation zur nächsten wird der Kampf um Ressourcen (den Darwin den „Kampf ums Dasein“ nannte) Individuen mit bestimmten Variationen gegenüber anderen bevorzugen und dadurch die Häufigkeit von Merkmalen innerhalb der Population verändern. Dieser Prozess ist die natürliche Selektion. Die Merkmale, die denjenigen Individuen einen Vorteil verschaffen, die mehr Nachkommen hinterlassen, werden als Anpassungen bezeichnet.
Damit die natürliche Auslese auf ein Merkmal einwirken kann, muss das Merkmal eine vererbbare Variation aufweisen und einen Vorteil im Wettbewerb um die Ressourcen verschaffen. Wenn eine dieser Voraussetzungen nicht gegeben ist, unterliegt das Merkmal nicht der natürlichen Selektion. (Wir wissen heute, dass sich solche Merkmale durch andere evolutionäre Mechanismen verändern können, die seit Darwins Zeit entdeckt wurden.)
Natürliche Auslese funktioniert durch komparative Vorteile, nicht durch einen absoluten Standard des Designs. „…da die natürliche Auslese durch den Wettbewerb um Ressourcen wirkt, passt sie die Bewohner eines jeden Landes nur im Verhältnis zum Grad der Vollkommenheit ihrer Mitbewohner an“ (Charles Darwin, Über die Entstehung der Arten, 1859).
Im zwanzigsten Jahrhundert wurde die Genetik in Darwins Mechanismus integriert, was uns erlaubt, die natürliche Auslese als das unterschiedliche Überleben und die Reproduktion von Genotypen zu bewerten, die bestimmten Phänotypen entsprechen. Die natürliche Auslese kann nur auf bestehende Variationen innerhalb einer Population einwirken. Solche Variationen entstehen durch Mutationen, d. h. durch Veränderungen in einem Teil des genetischen Codes für ein Merkmal. Mutationen entstehen zufällig und ohne Voraussicht auf den möglichen Vor- oder Nachteil der Mutation. Mit anderen Worten: Variationen entstehen nicht, weil sie gebraucht werden.
Beweise für die natürliche Auslese
Lassen Sie uns ein Beispiel betrachten, um die natürliche Auslese zu verdeutlichen.
Der industrielle Melanismus ist ein Phänomen, von dem über 70 Mottenarten in England betroffen sind. Es wurde am besten an der Pfeffermotte, Biston betularia, untersucht. Vor 1800 hatte der typische Falter dieser Art ein helles Muster (siehe Abbildung 2). Dunkel gefärbte oder melanische Falter waren selten und daher Sammlerstücke.

Abbildung 2. Abbildung 2: Pfeffermotte
Während der industriellen Revolution verdunkelten Ruß und andere Industrieabfälle die Baumstämme und töteten Flechten ab. Die helle Form des Nachtfalters wurde selten und die dunkle Form wurde immer häufiger. Im Jahr 1819 wurde die erste melanische Morphe gesichtet; bis 1886 war sie weitaus häufiger anzutreffen – ein Beleg für den raschen evolutionären Wandel. 
Schließlich waren helle Morphen nur noch an wenigen Orten weitab von Industriegebieten verbreitet. Als Ursache für diesen Wandel wurde die selektive Prädation durch Vögel vermutet, die die Tarnfärbung der Motte begünstigten.
In den 1950er Jahren führte der Biologe Kettlewell Wiederfangexperimente mit beiden Morphen durch. Eine kurze Zusammenfassung seiner Ergebnisse finden Sie unten. Durch die Beobachtung von Vögeln aus Jalousien konnte er bestätigen, dass die Auffälligkeit der Motte die Wahrscheinlichkeit, dass sie gefressen wird, stark beeinflusst.
Wiederfangerfolg
|
heller Falter |
dunkler Falter |
|
|
nichtindustrielle Wälder |
14.6 % |
4.7 % |
|
industrielle Hölzer |
13 % |
27.5 % |
Lokale Anpassung – weitere Beispiele
Bislang haben wir in der heutigen Vorlesung betont, dass die natürliche Selektion der Eckpfeiler der Evolutionstheorie ist. Sie liefert den Mechanismus für adaptive Veränderungen. Jede Veränderung in der Umwelt (z. B. eine Veränderung der Hintergrundfarbe des Baumstamms, auf dem man sich niederlässt) wird wahrscheinlich zu einer lokalen Anpassung führen. Jede weit verbreitete Population wird wahrscheinlich in verschiedenen Teilen ihres Verbreitungsgebiets unterschiedliche Umweltbedingungen vorfinden. Infolgedessen wird sie bald aus einer Reihe von Teilpopulationen bestehen, die sich geringfügig oder sogar erheblich unterscheiden.
Die folgenden Beispiele veranschaulichen die Anpassung von Populationen an lokale Bedingungen.
- Die Ringelnatter, Elaphe obsoleta, hat erkennbar unterschiedliche Populationen in verschiedenen Gegenden des östlichen Nordamerikas (siehe Abbildung 3). Ob diese als geografische „Rassen“ oder Unterarten bezeichnet werden sollten, ist umstritten. Diese Populationen bilden alle eine Art, da es zu Paarungen zwischen benachbarten Populationen kommen kann, wodurch die Arten einen gemeinsamen Genpool teilen (siehe die Vorlesung über Artbildung).
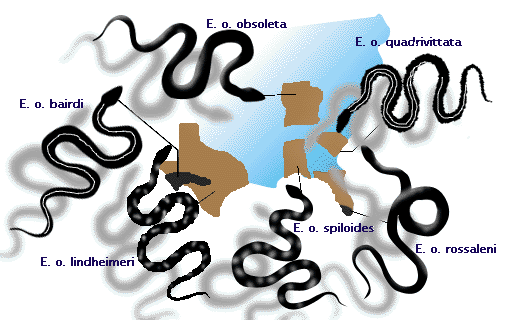
Abbildung 3: Unterarten der Ringelnatter Elaphe obsoleta, die sich dort kreuzen, wo sich ihre Verbreitungsgebiete treffen.
- Galapagosfinken sind das berühmte Beispiel von Darwins Reise. Jede Insel der Galapagos-Inseln, die Darwin besuchte, hatte ihre eigene Finkenart (insgesamt 14), die nirgendwo sonst auf der Welt vorkommt. Einige hatten Schnäbel, die für das Fressen großer Samen geeignet waren, andere für kleine Samen, einige hatten papageienähnliche Schnäbel zum Fressen von Knospen und Früchten, und einige hatten schlanke Schnäbel zum Fressen kleiner Insekten (siehe Abbildung 4). Einer benutzte einen Dorn, um nach Insektenlarven im Holz zu suchen, wie es einige Spechte tun. (Sechs waren Bodenbewohner und acht waren Baumfinken.) (Man nimmt an, dass diese Diversifizierung in verschiedene ökologische Rollen oder Nischen notwendig ist, um die Koexistenz mehrerer Arten zu ermöglichen, ein Thema, das wir in einer späteren Vorlesung untersuchen werden.) Für Darwin sah es so aus, als sei jede Art leicht verändert von einem ursprünglichen Kolonisten, wahrscheinlich dem Finken auf dem südamerikanischen Festland, etwa 600 Meilen östlich. Es ist wahrscheinlich, dass die adaptive Radiation zur Entstehung so vieler Arten führte, weil es nur wenige oder gar keine anderen Vögel gab, so dass leere Nischen zu besetzen waren, und weil die zahlreichen Inseln der Galapagos-Inseln reichlich Gelegenheit zur geografischen Isolation boten.
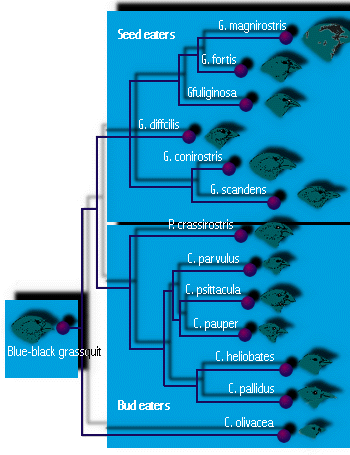
Abbildung 4
Stabilisierende, gerichtete und diversifizierende Selektion
Abschließend werden wir uns eine statistische Betrachtungsweise der Selektion ansehen. Nehmen wir an, dass jede Population als Häufigkeitsverteilung für ein Merkmal dargestellt werden kann – zum Beispiel die Schnabelgröße. Auch hier gilt, dass die Variation eines Merkmals das entscheidende Rohmaterial für die Evolution ist.
Wie wird die Häufigkeitsverteilung in der nächsten Generation aussehen?
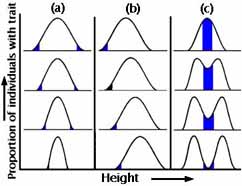
Abbildungen 5a-c
Erstens könnte der Anteil der Individuen mit jedem Wert des Merkmals (Größe des Schnabels oder Körpergewicht) genau gleich sein. Zweitens kann es zu einer Veränderung in nur eine Richtung kommen. Drittens (und das ist so selten, dass man darüber streiten kann) kann es zu gleichzeitigen Veränderungen in beide Richtungen kommen (z. B. werden sowohl größere als auch kleinere Schnäbel bevorzugt, auf Kosten derjenigen mit mittlerer Größe). Die Abbildungen 5a-c zeigen diese drei Hauptkategorien der natürlichen Selektion.
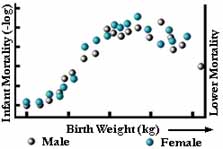
Abbildung 6
Bei der stabilisierenden Selektion werden extreme Varianten von beiden Enden der Häufigkeitsverteilung eliminiert. Die Häufigkeitsverteilung sieht genauso aus wie in der Generation davor (siehe Abbildung 5a). Dies ist wahrscheinlich die häufigste Form der natürlichen Selektion, und wir verwechseln sie oft mit keiner Selektion. Ein Beispiel aus dem wirklichen Leben ist das Geburtsgewicht von menschlichen Babys (siehe Abbildung 6).
Bei gerichteter Selektion schneiden Individuen an einem Ende der Verteilung der Schnabelgröße besonders gut ab, so dass die Häufigkeitsverteilung des Merkmals in der nachfolgenden Generation gegenüber derjenigen in der Elterngeneration verschoben ist (siehe Abbildung 5b). Dies ist das, was wir normalerweise als natürliche Selektion bezeichnen. Der industrielle Melanismus war ein solches Beispiel.

Abbildung 7
Der fossile Stammbaum des Pferdes liefert ein bemerkenswertes Beispiel für die gerichtete Sukzession. Der gesamte Stammbaum ist recht kompliziert und besteht nicht nur aus einer einfachen Linie vom winzigen Urpferd Hyracotherium aus dem frühen Eozän bis zum heute bekannten Equus. Im Großen und Ganzen hat sich das Pferd jedoch von einem kleinwüchsigen Vorfahren, der sich durch Wälder und Dickichte bewegte, zu einem langbeinigen Nachfahren entwickelt, der für Schnelligkeit auf offenem Grasland gebaut wurde. Diese Entwicklung war mit gut dokumentierten Veränderungen der Zähne, der Beinlänge und der Zehenstruktur verbunden (siehe Abbildung 7).
Bei einer diversifizierenden (disruptiven) Selektion werden beide Extreme auf Kosten von Zwischenformen bevorzugt (siehe Abbildung 5c). Dies ist ungewöhnlich, aber von theoretischem Interesse, weil es einen Mechanismus zur Artbildung ohne geographische Isolation nahelegt (siehe die Vorlesung über Artbildung).
Zusammenfassung
Darwins Evolutionstheorie änderte die Richtung des zukünftigen wissenschaftlichen Denkens grundlegend, obwohl sie auf einem wachsenden Korpus von Gedanken aufbaute, die begannen, frühere Vorstellungen über die natürliche Welt zu hinterfragen.
Der Kern von Darwins Theorie ist die natürliche Auslese, ein Prozess, der über aufeinanderfolgende Generationen abläuft und als unterschiedliche Reproduktion von Genotypen definiert wird.
Die natürliche Auslese erfordert eine vererbbare Variation eines bestimmten Merkmals und ein unterschiedliches Überleben und eine unterschiedliche Reproduktion, die mit dem Besitz dieses Merkmals verbunden sind.
Beispiele für natürliche Selektion sind gut dokumentiert, sowohl durch Beobachtung als auch durch den Fossilienbestand.
Die Selektion wirkt auf die Häufigkeit von Merkmalen und kann die Form einer stabilisierenden, gerichteten oder diversifizierenden Selektion annehmen.
Empfohlene Lektüre
o Darwin, C. 1959. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life. London: J. Murray.
o Futuyma, D.J. 1986. Evolutionary Biology. Sunderland, Mass: Sinauer Associates, Inc.
o Dawkins, R. 1989. The Selfish Gene. Oxford: Oxford University Press.