Introduction
In einer früheren Arbeit unseres Labors (Shalom, 2009) wurde argumentiert, dass die neurokognitive Verarbeitung in drei Ebenen unterteilt werden kann: (1) eine Basisebene mit primärer kognitiver, emotionaler und sensomotorischer Verarbeitung. So kann beispielsweise ein lautes, unerwartetes Geräusch, das über die primären auditorisch-sensorischen Systeme wahrgenommen wird, physiologisch-emotionale Reaktionen auslösen, wie z. B. Angst (schneller Herzschlag usw.). (2) eine integrative Ebene, die den Output aller primären Prozesse der Basisebene kombiniert und eine globale, kohärente Bedeutung, Erfahrung oder ein Verhalten bildet. Zum Beispiel führen die mentalen Repräsentationen der verschiedenen primären Elemente, die die emotionale Reaktion Angst ausmachen, zu einem bewussten Gefühl der Angst. (3) Eine logische Ebene, die aus der Basisebene abstrakte logische Regeln (Wenn-dann-Regeln) bildet (z. B. „Wenn ich schnelles Herzklopfen und kalten Schweiß habe, könnte ich Angst haben“) und auch an der Selektion und Hemmung beteiligt ist (z. B. Kontrolle emotionaler Triebe). Diese dreistufige Architektur wurde auf vier allgemeine psychologische Bereiche angewandt: Emotion, Gedächtnis, Empfindung-Wahrnehmung und Motorik. Shalom (2009) konzentrierte sich auf die zweite, integrative Ebene und ihre Beziehung zu den vier psychologischen Bereichen. Er argumentierte auch, dass diese vier Arten der Integration von vier verschiedenen Unterbereichen des medialen präfrontalen Kortex bedient werden: Brodmann Area (BA) 11 (Wahrnehmung), BA 10 (Gedächtnis), BA 9 (Emotion) und BA 8 (Motorik). Schließlich wurden Beweise dafür vorgelegt, dass ein selektives Defizit in diesen BA-Bereichen und diese Arten von integrativen Prozessen einigen der gemeinsamen Defizite bei ASD (Autismus-Spektrum-Störungen) zugrunde liegen.
Die aktuelle Übersichtsarbeit versucht, eine ähnliche Analyse für den lateralen präfrontalen Kortex (LPFC) durchzuführen, indem sie von den relevanten medialen präfrontalen Arealen auf ihre lateralen Gegenstücke übergeht: laterale BA 11/BA 47 (Wahrnehmung), laterale BA 10/BA 46 (Gedächtnis), laterale BA 9 (Emotion) und laterale BA 8 (Motorik) (Abbildung 1), (vgl. die Unterscheidung zwischen den Clustern 1, 2, 3 und 4 des anterioren Cingulums in Beckmann et al. (2009). Es gibt Hinweise darauf, dass eine selektive Atypizität in diesen BA-Arealen und ein selektives Defizit bei Selektions-/Hemmungsprozessen in diesen vier kognitiven Bereichen an einigen der häufigen Defizite bei ADHS (Aufmerksamkeitsdefizit-Hyperaktivitätsstörung) beteiligt ist, wie etwa Schwierigkeiten bei der Hemmung motorischer Reaktionen (z. B., Unfähigkeit, unangemessene Bewegungen zu hemmen), Wahrnehmungsfokus (z.B. Unfähigkeit, Ablenkungen zu ignorieren) und emotionale Reaktionen (z.B. Unfähigkeit, Triebe zu kontrollieren) (Barkley, 1997; Nigg, 1999).
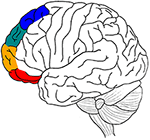
Abbildung 1. Eine grafische Zusammenfassung der wichtigsten anatomischen Hypothesen des Artikels. Motorische Verarbeitung in blau, Emotion in grün, Gedächtnis in orange und sensorisch-perzeptive Verarbeitung in rot.
Wahrnehmung
Wahrnehmungsselektion und -hemmung und laterale BA 11/47
Es gibt Hinweise darauf, dass Wahrnehmungsselektion und -hemmung durch neuronale Netzwerke unterstützt werden, die den lateralen Teil von BA 11/47 im LPFC einbeziehen. In einem von Zsuga et al. (2016) vorgeschlagenen Modell wird angenommen, dass der orbitofrontale Kortex (OFC) eine wichtige Rolle bei der Auswahl visueller Stimuli entsprechend den Aufgabenanforderungen spielt, wobei der mediale Teil des OFC eine zentrale Rolle bei der Integration und Kategorisierung von Informationen über Stimuli aus der Umgebung und ihrem Kontext spielt, während der laterale Teil des OFC an der Zuweisung und Aktualisierung von Auswahlparametern entsprechend den aufgabenspezifischen Werten der verfügbaren Stimuli beteiligt ist. Zur Unterstützung dieser Behauptung zeigten Howard und Kahnt (2017) in einer Studie, dass der laterale Teil des BA 11 an der Kodierung zielgerichteter Werte von Geruchsreizen beteiligt ist. Sie präsentierten hungrigen Teilnehmern zwei bevorzugte Essensgerüche und forderten sie auf, zu entscheiden, ob sie den einen oder den anderen riechen wollten. Später bekamen die Teilnehmer eines der beiden Lebensmittel zu essen und durften erneut zwischen den beiden Gerüchen wählen. Die Veränderung der Präferenz für den nicht gesättigten Geruch spiegelte sich in einer Veränderung der Aktivität im lateralen Teil von BA 11/47 wider. Darüber hinaus wurde gezeigt, dass der laterale BA 11/47 an der Ablehnung irrelevanter Reize beteiligt ist (Kaufman et al., 2016).
Der laterale BA 11/47 ist nachweislich auch an der Speicherung visueller Reize im Arbeitsgedächtnis und der Steuerung der visuellen Auswahl beteiligt. In einer Studie verglichen Soto et al. (2007) zwei Bedingungen für eine visuelle Auswahl, bei der die Grundierung entweder einmal oder zweimal gezeigt wurde. Die laterale BA 11/47-Aktivierung war größer, wenn der Primer zweimal gezeigt wurde, jedoch wurde eine Verringerung der Aktivierung festgestellt, wenn ähnliche Reize zweimal passiv wiederholt wurden (ohne visuelle Auswahlphase und daher ohne Aufgabenrelevanz).
Schließlich ist laut Price (2007) die laterale BA 11 Teil des sensorischen orbitalen Netzwerks.
ADHS und visuelle Selektion und Hemmung
Es gibt Hinweise darauf, dass Kinder und Erwachsene mit ADHS Probleme mit der Hemmung irrelevanter visueller und auditorischer Reize haben. So zeigten Erwachsene mit ADHS beispielsweise mehr Interferenzeffekte (d. h. Probleme bei der Hemmung irrelevanter Reize) als Kontrollpersonen, wenn sie eine Erinnerungsaufgabe lösen sollten, während sie ein Hintergrundgeräusch ignorierten (Pelletier et al., 2016). Darüber hinaus wurde eine positive Korrelation zwischen der Aufgabenleistung unter irrelevanten Geräuschen und dem Ausmaß der von den Patienten auf einer klinischen Symptomskala angegebenen Aufmerksamkeitssymptome festgestellt. In einer anderen Studie wurden Erwachsene mit ADHS und Kontrollpersonen gebeten, während des Autofahrens ein Telefongespräch zu führen. Die ADHS-Gruppe zeigte im Vergleich zu den Kontrollpersonen während des Telefongesprächs signifikant schlechtere Fahrfähigkeiten als in der Ruhephase (Reimer et al., 2010). Andere Studien haben gezeigt, dass die visuelle Hemmung bei ADHS-Personen schlecht ist. So fanden Forster et al. (2014) und Forster und Lavie (2016) heraus, dass Erwachsene mit ADHS eine Buchstabensuchaufgabe langsamer ausführten, wenn irrelevante, aber auffällige Zeichentrickfiguren auf dem Bildschirm zu sehen waren, und zwar in höherem Maße als Kontrollpersonen. In ähnlicher Weise erfordert die Stroop-Interferenzaufgabe, dass die Teilnehmer eine bestimmte Dimension eines visuellen Reizes beachten, während sie eine andere ignorieren. In Studien wurde festgestellt, dass ADHS-Kinder bei der Stroop-Interferenz-Aufgabe schlechter abschneiden als Kontrollpersonen (Sørensen et al., 2014) und dass ihre Leistung mit ihren Unaufmerksamkeits- und Hyperaktivitätssymptomen korreliert (Ikeda et al., 2013).
ADHS, visuelle Selektion und Hemmung und laterale BA 11/47
Außerdem fanden mehrere Studien spezifische Hinweise auf eine atypische Aktivierung in der lateralen BA 11/47 bei Personen mit ADHS, die visuelle Selektions-/Hemmungsaufgaben durchführen. Tsujimoto et al. (2013) baten Kinder mit und ohne ADHS, eine Arbeitsgedächtnisaufgabe mit und ohne Ablenkung durchzuführen. Die ADHS-Teilnehmer zeigten signifikant schlechtere Verhaltensleistungen, insbesondere unter Ablenkung, und wiesen signifikant höhere Aktivierungswerte im lateralen BA 11/47 auf als die Kontrollpersonen. In einer anderen Studie testeten Yasumura et al. (2014) Kinder mit ADHS und Kontrollkinder sowohl unter Stroop-Interferenz- als auch unter umgekehrten Stroop-Interferenzbedingungen und stellten fest, dass ADHS-Kinder unter beiden Bedingungen schlechtere Leistungen erbrachten und dass die Leistung mit der Aktivität der lateralen BA 47 korrelierte. In einer anderen Studie, in der ein auffälliger, aber irrelevanter Distraktor während einer Zeitschätzungsaufgabe präsentiert wurde, wurde festgestellt, dass der Distraktor den ADHS-Teilnehmern zwar im gleichen Maße half wie den Kontrollpersonen, die laterale BA 11-Aktivierung, die mit dem Auftauchen des Distraktors einherging, jedoch deutlich größer war (Pretus et al, 2016).
Gedächtnis
Gedächtnisselektion und -hemmung und laterale BA 10/46
Es besteht zwar kein allgemeiner Konsens darüber, dass BA10/46 an Funktionen des episodischen Gedächtnisses beteiligt ist, aber es gibt viele Belege für eine Rolle von BA 10/46 bei der Auswahl und Hemmung von Gedächtnisinformationen. Kim (2011) analysierte beispielsweise 74 Studien, die das Erinnern und Vergessen verglichen (d. h. erfolgreich erinnerte Elemente im Vergleich zu fälschlicherweise als erschienen bewerteten Distraktoren), und stellte fest, dass sich die meisten Studien im frontalen Teil von BA 46 überschnitten, in der Nähe der Verbindung mit dem lateralen BA 10. In einer späteren Studie führte Kim (2013) eine Meta-Analyse von 70 Studien durch, die sich auf neue und alte Items konzentrierten (d.h. Treffer vs. korrekte Ablehnungen). Die Ergebnisse zeigen, dass neue>alte Vergleiche zu Aktivierungen führen, die sich über den mittleren Teil des linken lateralen präfrontalen Bereichs ausbreiten. Wenn man sich jedoch auf Studien zum Quellgedächtnis konzentriert, bei denen die Teilnehmer nicht nur das Zielobjekt, sondern auch ein zusätzliches Detail aus der Lernphase abrufen müssen (d. h., (d.h. wenn die Selektionsanforderungen höher sind), wurde eine spezifische Aktivierung im frontalen Teil von BA46, einschließlich des rostralen Teils von BA10, beobachtet.
Eine andere Linie von Beweisen stammt aus einer Meta-Analyse von Gilbert et al. (2006), in der 104 Studien mit Schwerpunkt auf BA10 analysiert wurden, und ergab, dass die Mehrheit der Studien, die den lateralen Teil von BA10 einbeziehen, das Arbeitsgedächtnis und den Abruf des episodischen Gedächtnisses testeten, während Studien, die den medialen Teil von BA10 einbeziehen, das Mentalisieren (d.h.,
Zusätzlich verwendeten Cohen et al. (2014) die Aufgabe des wertorientierten Erinnerns, bei der den Teilnehmern Wortlisten vorgelegt werden, in denen einigen Wörtern ein höherer Wert als anderen zugewiesen wird, was die Teilnehmer dazu anregt, während der Kodierung höherwertige Elemente gegenüber geringwertigen auszuwählen. Die Ergebnisse zeigen eine spezifische Aktivierung der rechten lateralen BA46/10 und eine sehr ausgeprägte Aktivierung der linken lateralen BA46/10, wenn ein Hinweis auf ein höherwertiges Wort erschien.
ADHS und Gedächtnisauswahl und -hemmung
Es gibt mehrere Studien, die Hinweise auf Schwierigkeiten bei der Auswahl und Hemmung von Gedächtnisinformationen bei ADHS gefunden haben. In einer Studie von Pollak et al. (2007) beispielsweise führten Erwachsene mit und ohne ADHS eine schwierige Listenlernaufgabe durch, bei der sie sich achtmal fünf Wortlisten merken und abrufen mussten. Die Ergebnisse zeigen, dass die ADHS-Teilnehmer mehr Doppelabrufe und Intrusionsfehler (Abruf von Wörtern aus der falschen Liste) machten, was auf eine Schwierigkeit bei der Hemmung des Abrufs von für die Aufgabe irrelevanten Elementen hinweist. In einer ähnlichen Studie baten Soliman und Elfar (2017) Erwachsene mit und ohne ADHS, acht Listen zu lernen, und stellten ihnen dann eine Erkennungsaufgabe. Die ADHS-Gruppe erkannte weniger Wörter richtig, gab mehr falsch-positive Antworten (d. h. sie wählte fälschlicherweise Elemente aus, die nicht gelernt worden waren) und war sich ihrer Fehler sicherer. In einer anderen Studie baten Castel et al. (2011) Kinder mit und ohne ADHS, das oben erwähnte wertorientierte Erinnerungsparadigma durchzuführen. Die Ergebnisse zeigen, dass sich Kinder mit ADHS an genauso viele Gegenstände erinnern konnten wie die Kontrollpersonen, aber sie waren weniger in der Lage, die Gegenstände auszuwählen, an die sie sich erinnern wollten.
ADHS, Gedächtnisauswahl und -hemmung und laterale BA 10/46
Die Literaturrecherche ergab nur eine Neuroimaging-Studie, die die Auswahl oder Hemmung von Gedächtnisfunktionen bei ADHS untersuchte. In einer Studie von Depue et al. (2010) wurde Erwachsenen mit und ohne ADHS eine Beziehung zwischen Gesicht-Bild-Paaren beigebracht, bis sie diese beherrschten. Später wurde ihnen ein Bild eines Gesichts gezeigt, und sie wurden aufgefordert, entweder an das zugehörige Bild zu denken oder zu verhindern, dass das zugehörige Bild ins Bewusstsein kommt. Die Ergebnisse zeigen, dass die ADHS-Teilnehmer die laterale BA 10 weniger aktivierten als die Kontrollpersonen, wenn man die „Nicht-Denken“-Bedingung mit der „Denken“-Bedingung verglich.
Emotion
Emotionsauswahl und -hemmung und laterale BA 9
Es gibt viele Belege für den Zusammenhang zwischen Emotionsregulation (die Hemmung bestimmter Emotionen und die Auswahl anderer) und lateraler BA 9. In einer Studie von Hallam et al. (2014) beispielsweise sahen die Teilnehmer emotionsauslösende Filme und wurden aufgefordert, ihre emotionale Reaktion entweder zu unterdrücken oder neu zu bewerten oder den Film einfach nur anzusehen. Die Ergebnisse zeigten, dass sowohl bei der Aufarbeitung als auch bei der Unterdrückung im Vergleich zur passiven Bedingung eine Aktivierung der lateralen BA 9 auftrat. In ähnlicher Weise wurde in zwei Studien die Aufarbeitung und das passive Betrachten von Bildern bei Menschen mit PTBS und Kontrollpersonen verglichen. In beiden Studien wurde festgestellt, dass die laterale BA 9 während der Reappraisal-Bedingung stärker aktiviert wurde, allerdings in geringerem Ausmaß bei Personen mit PTBS (Xiong et al., 2013; Rabinak et al., 2014).
Zusätzlich wurden zwei verschiedene Meta-Analysen verwendet, um fMRI-Studien zur Bewertung der Emotionsregulation zu untersuchen. Buhle et al. (2014) fanden heraus, dass eine Aufarbeitung, die eine Herabregulierung des negativen Affekts beinhaltet, durchgängig die laterale BA 9 aktivierte; Frank et al. (2014) fanden heraus, dass eine solche Aufarbeitung mit einer stärkeren Aktivierung in der lateralen BA 9 und einer geringeren Aktivierung in der Amygdala einherging.
ADHS und Emotionsselektion und -hemmung
Verschiedene Studien fanden Defizite in der Emotionsregulation bei Kindern mit ADHS. So zeigen mehrere Studien, dass es Kindern mit ADHS schwerer fällt, ihre Emotionen zu unterdrücken als Gleichaltrigen mit typischer Entwicklung (TD). So wurden in einer Studie Kinder einem Wettbewerb unter Gleichaltrigen unterzogen, bei dem die Hälfte der Kinder ihre Emotionen verbergen sollte und die andere Hälfte nicht. Die Ergebnisse zeigen, dass ADHS-Kinder ihre Emotionen weniger gut verbergen konnten als ihre Altersgenossen (Walcott und Landau, 2004). Darüber hinaus ergab eine Studie, die die Leistung von Kindern mit ADHS bei einer emotionalen Stroop-Aufgabe untersuchte, dass es Kindern mit ADHS schwerer fiel, Reaktionen auf wütende und frustrierte Gesichter zu hemmen (Yarmolovsky et al., 2016).
In einer von Graziano und Garcia (2016) durchgeführten Metaanalyse zur Emotionsregulation bei ADHS fanden sie außerdem vier Bereiche von Interesse, nämlich: Erkennen (d. h., Erkennen (d. h. die Fähigkeit, die Emotionen anderer und der eigenen Person zu verarbeiten und abzuleiten), Reaktivität (d. h. die Schwelle, Intensität und Dauer der eigenen affektiven Erregung), Regulierung (d. h. die effektive Reaktion auf emotionale Reaktivität in einer flexiblen Art und Weise, die das adaptive Funktionieren erleichtert) und Empathie (d. h. die Fähigkeit, den affektiven Zustand einer anderen Person zu erleben und/oder die Besorgnis über die Lage einer anderen Person auszudrücken). Die Ergebnisse zeigen, dass zwar alle vier Bereiche in der ADHS-Population atypisch zu sein scheinen, die auffälligsten Defizite jedoch in den Bereichen Reaktion und Regulation auftreten. Dies bedeutet, dass (insbesondere) Kinder mit ADHS dazu neigen, schneller, intensiver und länger auf aversive Situationen zu reagieren, und weniger in der Lage sind, diese Emotionen zu regulieren, selbst wenn sie dies versuchen.
ADHS, Emotionsselektion und Hemmung und laterale BA 9
Es gibt mehrere Studien, die spezifische Atypien in der lateralen BA 9-Aktivierung bei ADHS-Teilnehmern zeigen, die Aufgaben ausführen, die eine Emotionsregulation erfordern. In einer Studie von Passarotti et al. (2010a) zum Beispiel führten Kinder mit bipolarer Störung, Kinder mit ADHS und eine Gruppe altersgleicher Kontrollpersonen eine emotionale Valenz-Stroop-Aufgabe durch. Bei dieser Aufgabe wurden positive, neutrale und negative Wörter präsentiert, und die Teilnehmer mussten entsprechend der Farbe des Wortes reagieren, während sie dessen Bedeutung ignorieren sollten. Die Ergebnisse zeigen, dass die bipolare Gruppe Schwierigkeiten hatte, positive Wörter zu ignorieren, während die ADHS-Gruppe Schwierigkeiten hatte, negative Wörter zu ignorieren. Interessanterweise war die laterale BA 9-Aktivierung ausgeprägter bei Aufgaben, die erfolgreich gelöst wurden, d. h. bei negativen Wörtern in der bipolaren Gruppe und bei positiven Wörtern in der ADHS-Gruppe. Darüber hinaus baten Hwang et al. (2015) in einer auf Stroop basierenden emotionalen Aufgabe Kinder mit und ohne ADHS, eine numerische Stroop-Aufgabe durchzuführen, nachdem ihnen Bilder gezeigt wurden, die entweder positiv, neutral oder negativ waren. Während die Verhaltensdaten keine signifikanten Unterschiede zwischen den verschiedenen emotionalen Bedingungen zeigten, stellten sie fest, dass Kinder mit ADHS weniger in der Lage waren, die laterale BA 9 zu rekrutieren, wenn sie die Aufgabe durchführten, ein Merkmal, das auch mit der Symptomschwere korrelierte, die mit der Conner-Eltern-Skala gemessen wurde.
Motorik
Motorische Auswahl und Hemmung und laterale BA 8
Es gibt viele Hinweise darauf, dass die Hemmung motorischer Handlungen durch neuronale Netzwerke in der lateralen BA 8 unterstützt wird. Die gebräuchlichste Messung der motorischen Inhibition ist die Stop-Signal-Aufgabe (SST), bei der die Teilnehmer aufgefordert werden, eine Taste zu drücken, wenn ein Stimulus präsentiert wird, diesen Tastendruck aber nicht auszuführen, wenn dem Stimulus kurz darauf ein Signal folgt. Studien zeigen, dass die Hemmung des Tastendrucks mit einer Aktivierung der lateralen BA 8 einhergeht (Matthews et al., 2005; Smith et al., 2013; Hughes et al., 2014). Es wurde auch gezeigt, dass die laterale BA 8 besonders aktiv ist, wenn die Auswahl einer motorischen Reaktion Aufmerksamkeit erfordert oder einen Konflikt beinhaltet. So forderten Enriquez-Geppert et al. (2013) die Teilnehmer auf, eine Taste zu drücken, wenn ein Stimulus auf dem Bildschirm erschien, wobei sich die Antworttaste in einigen Versuchen änderte. Die Experimentatoren fanden heraus, dass die laterale BA 8 speziell bei Versuchen aktiviert wurde, bei denen die Antworttaste anders war. In ähnlicher Weise verwendeten Lenartowicz et al. (2011) eine Go/No-Go-Aufgabe, bei der sich die Stimuli nach einigen Übungssitzungen änderten, so dass aus dem vorherigen Go-Signal ein No-Go-Signal wurde (was eine stärkere Hemmung erforderte). Sie stellten fest, dass die laterale BA 8 bei diesen Versuchen spezifisch aktiviert wurde. In einer ähnlichen Studie verwendeten Albares et al. (2014) eine Version einer Go/No-Go-Aufgabe, bei der vor den Go- und No-Go-Reizen eine Prime präsentiert wurde. Während jedoch ein grünes Licht anzeigte, dass ein Go-Reiz folgen würde, bedeutete ein rotes Licht, dass eine von drei Bedingungen eintreten konnte: entweder ein Go-Reiz, ein No-Go-Reiz oder gar kein Reiz. Die Forscher schlussfolgerten, dass die Rotlicht-Bedingung eine komplexere motorische Planung erfordert als die Grünlicht-Bedingung, und fanden heraus, dass die laterale BA 8 während dieser Bedingung tatsächlich aktiver war.
ADHS und motorische Selektion und Hemmung
Es gibt deutliche Hinweise auf Defizite bei der motorischen Selektion und Hemmung bei Kindern mit ADHS. Viele bisherige Studien haben gezeigt, dass Kinder, Jugendliche und Erwachsene mit ADHS bei verschiedenen Versionen der Stoppsignalaufgabe schlechtere Leistungen zeigen (z. B. Rubia et al., 1998; Lee et al., 2016; Bialystok et al., 2017; Dekkers et al., 2017), und dass diese Unterschiede verschwinden können, wenn Kinder mit ADHS angemessen medikamentös behandelt und durch effektive Verstärkung motiviert werden (Rosch et al., 2016). Darüber hinaus wurden in einer sehr großen Studie von Crosbie et al. (2013) ADHS-Symptome bei über 16 000 Kindern aus der Allgemeinheit gemessen, die den SST durchführten. Die Ergebnisse zeigten, dass Personen mit höheren ADHS-Merkmalswerten eine schlechtere Reaktionshemmung, eine langsamere Reaktionslatenz und eine größere Variabilität aufwiesen und dass dieser Trend in hohem Maße vererbbar war. In ähnlicher Weise führten Alderson et al. (2007) eine Meta-Analyse durch, die sich auf ADHS-Kinder und den SST konzentrierte, und fanden in der ADHS-Gruppe größere mittlere Antwortzeiten, eine größere Antwortvariabilität und eine größere Stoppsignal-Antwortzeit.
ADHS, motorische Selektion und Hemmung und laterale BA 8
Es gibt auch beträchtliche Belege für spezifische Atypizitäten in der lateralen BA 8-Aktivierung bei ADHS-Teilnehmern, die Aufgaben ausführen, die motorische Selektion und Hemmung erfordern. So wurden beispielsweise in zwei separaten Meta-Analysen fMRT-Studien zur Bewertung der motorischen Hemmung bei Kindern und Erwachsenen mit ADHS untersucht (Hart et al., 2013; Rubia et al., 2013). Beide Meta-Analysen ergaben, dass eine geringere Aktivierung in der lateralen BA 8 mit der Messung der motorischen Hemmung zusammenhängt, und in der Studie von Rubia et al. (2013) wurde auch festgestellt, dass Methylphenidat die Aktivierung in der lateralen BA 8 erhöht, was mit einer verbesserten Leistung bei motorischen Aufgaben einhergeht. Darüber hinaus wurde in Studien, in denen verschiedene Versionen des SST und der Go/No-Go-Aufgabe verwendet wurden, eine geringere Aktivierung der lateralen BA 8 in der ADHS-Gruppe während einer einfachen No-Go-Bedingung (Passarotti et al., 2010b; Mulligan et al., 2011), kurz vor dem Auftreten eines Fehlers (Spinelli et al., 2011) sowie während einer Free-Choice-Bedingung festgestellt, bei der die Teilnehmer entschieden, ob sie einen Knopf drücken wollten oder nicht, wenn ein Stimulus präsentiert wurde (Karch et al., 2010).
Zusammenfassung
Die vorliegende Arbeit präsentiert Belege dafür, dass der laterale Teil des präfrontalen Kortex an der Hemmung, Auswahl und Manipulation motorischer, emotionaler, gedächtnisbezogener und wahrnehmungssensorischer Informationen beteiligt ist, eine Funktion, die im Alltag hilfreich ist, da sie es uns ermöglicht, Ablenkungen zu ignorieren, spezifische Informationen aus unserem Gedächtnis auszuwählen und abzurufen, unseren emotionalen Zustand zu erkennen und zu regulieren oder eine situationsgerechte motorische Reaktion zu planen. Anhand dieses Modells werden die Probleme von Menschen mit ADHS klassifiziert, wobei verschiedene Arten von Schwierigkeiten bei der Selektion/Hemmung mit verschiedenen Teilbereichen des lateralen präfrontalen Kortex in Verbindung gebracht werden.
Autorenbeiträge
Der Autor bestätigt, dass er der alleinige Verfasser dieser Arbeit ist und sie zur Veröffentlichung freigegeben hat.
Erklärung zu Interessenkonflikten
Der Autor erklärt, dass die Forschung in Abwesenheit von kommerziellen oder finanziellen Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Albares, M., Lio, G., Criaud, M., Anton, J. L., Desmurget, M., und Boulinguez, P. (2014). Der dorsale mediale frontale Kortex vermittelt die automatische motorische Hemmung in unsicheren Kontexten: Hinweise aus kombinierten fMRI- und EEG-Studien. Hum. Brain Mapp. 35, 5517-5531. doi: 10.1002/hbm.22567
PubMed Abstract | CrossRef Full Text | Google Scholar
Alderson, R. M., Rapport, M. D., and Kofler, M. J. (2007). Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung und Verhaltenshemmung: eine meta-analytische Überprüfung des Stop-Signal-Paradigmas. J. Abnorm. Child Psychol. 35, 745-758. doi: 10.1007/s10802-007-9131-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Barkley, R. A. (1997). Verhaltenshemmung, anhaltende Aufmerksamkeit und exekutive Funktionen: Konstruktion einer vereinheitlichenden Theorie der ADHS. Psychol. Bull. 121:65. doi: 10.1037/0033-2909.121.1.65
PubMed Abstract | CrossRef Full Text | Google Scholar
Beckmann, M., Johansen-Berg, H., and Rushworth, M. F. (2009). Konnektivitätsbasierte Parzellierung des menschlichen cingulären Cortex und seine Beziehung zur funktionellen Spezialisierung. J. Neurosci. 29, 1175-1190. doi: 10.1523/JNEUROSCI.3328-08.2009
PubMed Abstract | CrossRef Full Text | Google Scholar
Bialystok, E., Hawrylewicz, K., Wiseheart, M., and Toplak, M. (2017). Interaktion von Zweisprachigkeit und Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung bei jungen Erwachsenen. Bilingualism 20, 588-601. doi: 10.1017/S1366728915000887
PubMed Abstract | CrossRef Full Text | Google Scholar
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., et al. (2014). Kognitive Aufarbeitung von Emotionen: eine Meta-Analyse menschlicher Neuroimaging-Studien. Cereb. Cortex 24, 2981-2990. doi: 10.1093/cercor/bht154
PubMed Abstract | CrossRef Full Text | Google Scholar
Castel, A. D., Lee, S. S., Humphreys, K. L., and Moore, A. N. (2011). Gedächtniskapazität, selektive Kontrolle und wertorientiertes Erinnern bei Kindern mit und ohne Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung (ADHS). Neuropsychology 25:15. doi: 10.1037/a0020298
PubMed Abstract | CrossRef Full Text | Google Scholar
Cohen, M. S., Rissman, J., Suthana, N. A., Castel, A. D., and Knowlton, B. J. (2014). Wertbasierte Modulation der Gedächtniskodierung beinhaltet die strategische Einbindung von fronto-temporalen semantischen Verarbeitungsregionen. Cogn. Affect. Behav. Neurosci. 14, 578-592. doi: 10.3758/s13415-014-0275-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crosbie, J., Arnold, P., Paterson, A., Swanson, J., Dupuis, A., Li, X., et al. (2013). Reaktionshemmung und ADHS-Merkmale: Korrelate und Heritabilität in einer Gemeinschaftsstichprobe. J. Abnorm. Child Psychol. 41, 497-507. doi: 10.1007/s10802-012-9693-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Dekkers, T. J., van Rentergem, J. A. A., Koole, A., van den Wildenberg, W. P., Popma, A., Bexkens, A., et al. (2017). Time-on-task effects in children with and without ADHD: depletion of executive resources or depletion of motivation? Eur. Child Adolesc. Psychiatry. 26, 1-11. doi: 10.1007/s00787-017-1006-y
CrossRef Full Text | Google Scholar
Depue, B. E., Burgess, G. C., Willcutt, E. G., Ruzic, L., and Banich, M. T. (2010). Inhibitorische Kontrolle des Gedächtnisabrufs und der motorischen Verarbeitung in Verbindung mit dem rechten lateralen präfrontalen Kortex: Nachweis von Defiziten bei Personen mit ADHS. Neuropsychologia. 48, 3909-3917. doi: 10.1016/j.neuropsychologia.2010.09.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Enriquez-Geppert, S., Eichele, T., Specht, K., Kugel, H., Pantev, C., and Huster, R. J. (2013). Funktionelle Parzellierung des inferioren frontalen und midcingulären Kortex in einem Flanker-Stopp-Wechsel-Paradigma. Hum. Brain Mapp. 34, 1501-1514. doi: 10.1002/hbm.22002
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., and Lavie, N. (2016). Die Etablierung des Merkmals der Aufmerksamkeitsablenkbarkeit. Psychol. Sci. 27, 203-212. doi: 10.1177/0956797615617761
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., Robertson, D. J., Jennings, A., Asherson, P., and Lavie, N. (2014). Das Aufmerksamkeitsdefizit ausgleichen: Wahrnehmungsbelastung wirkt erhöhter Ablenkung bei ADHS entgegen. Neuropsychology 28:91. doi: 10.1037/neu0000020
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank, D. W., Dewitt, M., Hudgens-Haney, M., Schaeffer, D. J., Ball, B. H., Schwarz, N. F., et al. (2014). Emotionsregulation: quantitative Meta-Analyse der funktionellen Aktivierung und Deaktivierung. Neurosci. Biobehav. Rev. 45, 202-211. doi: 10.1016/j.neubiorev.2014.06.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Gilbert, S. J., Spengler, S., Simons, J. S., Steele, J. D., Lawrie, S. M., Frith, C. D., et al. (2006). Funktionelle Spezialisierung im rostralen präfrontalen Kortex (Area 10): eine Meta-Analyse. J. Cogn. Neurosci. 18, 932-948. doi: 10.1162/jocn.2006.18.6.932
PubMed Abstract | CrossRef Full Text | Google Scholar
Graziano, P. A., and Garcia, A. (2016). Aufmerksamkeitsdefizit-Hyperaktivitätsstörung und Emotionsdysregulation bei Kindern: eine Meta-Analyse. Clin. Psychol. Rev. 46, 106-123. doi: 10.1016/j.cpr.2016.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Hallam, G. P., Webb, T. L., Sheeran, P., Miles, E., Niven, K., Wilkinson, I. D., et al. (2014). Die neuronalen Korrelate der Regulierung der Emotionen einer anderen Person: eine explorative fMRI-Studie. Front. Hum. Neurosci. 8:376. doi: 10.3389/fnhum.2014.00376
PubMed Abstract | CrossRef Full Text | Google Scholar
Hart, H., Radua, J., Nakao, T., Mataix-Cols, D., and Rubia, K. (2013). Meta-Analyse von Studien zur funktionellen Magnetresonanztomographie über Hemmung und Aufmerksamkeit bei Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung: Untersuchung von aufgabenspezifischen, stimulierenden Medikamenten- und Alterseffekten. JAMA Psychiatry. 70, 185-198. doi: 10.1001/jamapsychiatry.2013.277
PubMed Abstract | CrossRef Full Text | Google Scholar
Howard, J. D., and Kahnt, T. (2017). Identitätsspezifische Belohnungsrepräsentationen im orbitofrontalen Kortex werden durch selektive Abwertung moduliert. J. Neurosci. 37, 3473-3416. doi: 10.1523/JNEUROSCI.3473-16.2017
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, M. E., Budd, T. W., Fulham, W. R., Lancaster, S., Woods, W., Rossell, S. L., et al. (2014). Anhaltende Hirnaktivierung zur Unterstützung der Leistung bei Stoppsignalen. Eur. J. Neurosci. 39, 1363-1369. doi: 10.1111/ejn.12497
PubMed Abstract | CrossRef Full Text | Google Scholar
Hwang, S., White, S. F., Nolan, Z. T., Craig Williams, W., Sinclair, S., and Blair, R. J. (2015). Exekutive Aufmerksamkeitskontrolle und emotionales Reagieren bei Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung – eine funktionelle MRT-Studie. Neuroimage Clin. 9, 545-554. doi: 10.1016/j.nicl.2015.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Ikeda, Y., Okuzumi, H., and Kokubun, M. (2013). Stroop/Reverse-Stroop-Interferenz in der typischen Entwicklung und ihre Beziehung zu Symptomen von ADHS. Res. Dev. Disabil. 34, 2391-2398. doi: 10.1016/j.ridd.2013.04.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Karch, S., Thalmeier, T., Lutz, J., Cerovecki, A., Opgen-Rhein, M., Hock, B., et al. (2010). Neuronale Korrelate (ERP/fMRI) der freiwilligen Auswahl bei erwachsenen ADHS-Patienten. Eur. Arch. Psychiatry Clin. Neurosci. 260, 427-440. doi: 10.1007/s00406-009-0089-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaufman, D. A., Keith, C. M., and Perlstein, W. M. (2016). Orbitofrontaler Kortex und die frühe Verarbeitung von visueller Neuheit im gesunden Altern. Front. Aging Neurosci. 8:101. doi: 10.3389/fnagi.2016.00101
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2011). Neuronale Aktivität, die späteres Erinnern und Vergessen vorhersagt: eine Meta-Analyse von 74 fMRI-Studien. Neuroimage 54, 2446-2461. doi: 10.1016/j.neuroimage.2010.09.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2013). Differentielle neuronale Aktivität bei der Erkennung von alten versus neuen Ereignissen: eine Aktivierungswahrscheinlichkeitsschätzungs-Meta-Analyse. Hum. Brain Mapp. 34, 814-836. doi: 10.1002/hbm.21474
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. Y., Wu, T. F., Tsai, J. D., and Yang, E. L. (2016). Anwendbarkeit der Stopp-Signal-Aufgabe für Vorschulkinder mit ADHS. Percept. Mot. Skills. 123, 162-174. doi: 10.1177/0031512516660715
PubMed Abstract | CrossRef Full Text | Google Scholar
Lenartowicz, A., Verbruggen, F., Logan, G. D., and Poldrack, R. A. (2011). Hemmungsbezogene Aktivierung im rechten inferioren frontalen Gyrus in Abwesenheit von hemmenden Hinweisen. J. Cogn. Neurosci. 23, 3388-3399. doi: 10.1162/jocn_a_00031
PubMed Abstract | CrossRef Full Text | Google Scholar
Matthews, S. C., Simmons, A. N., Arce, E., and Paulus, M. P. (2005). Dissoziation von Inhibition und Fehlerverarbeitung unter Verwendung einer parametrischen inhibitorischen Aufgabe während der funktionellen Magnetresonanztomographie. Neuroreport. 16, 755-760. doi: 10.1097/00001756-200505120-00020
PubMed Abstract | CrossRef Full Text | Google Scholar
Mulligan, R. C., Knopik, V. S., Sweet, L. H., Fischer, M., Seidenberg, M., and Rao, S. M. (2011). Neuronale Korrelate der hemmenden Kontrolle bei Erwachsenen mit Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung: Erkenntnisse aus der Milwaukee-Längsschnittstudie. Psychiatry Res. 194, 119-129. doi: 10.1016/j.pscychresns.2011.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Nigg, J. T. (1999). Das ADHS-Reaktionshemmungsdefizit, gemessen mit der Stopp-Aufgabe: Replikation mit DSM-IV kombiniertem Typ, Erweiterung und Qualifikation. J. Abnorm. Child Psychol. 27, 393-402. doi: 10.1023/A:1021980002473
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010a). Neuronale Korrelate der Reaktionshemmung bei pädiatrischer bipolarer Störung und Aufmerksamkeitsdefizit-Hyperaktivitätsstörung. Psychiatry Res. Neuroimaging. 181, 36-43. doi: 10.1016/j.pscychresns.2009.07.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010b). Unterschiedliche Beteiligung kognitiver und affektiver neuronaler Systeme bei pädiatrischer bipolarer Störung und Aufmerksamkeitsdefizit-Hyperaktivitätsstörung. J. Int. Neuropsychol. Soc. 16, 106-117. doi: 10.1017/S1355617709991019
PubMed Abstract | CrossRef Full Text | Google Scholar
Pelletier, M. F., Hodgetts, H. M., Lafleur, M. F., Vincent, A., and Tremblay, S. (2016). Anfälligkeit für den irrelevanten Geräuscheffekt bei ADHS bei Erwachsenen. J. Atten. Disord. 20, 306-316. doi: 10.1177/1087054713492563
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollak, Y., Kahana-Vax, G., and Hoofien, D. (2007). Abrufprozesse bei Erwachsenen mit ADHS: eine RAVLT-Studie. Dev. Neuropsychol. 33, 62-73. doi: 10.1080/87565640701729789
PubMed Abstract | CrossRef Full Text | Google Scholar
Pretus, C., Picado, M., Ramos-Quiroga, A., Carmona, S., Richarte, V., Fauquet, J., et al. (2016). Die Anwesenheit eines Distraktors verbessert die Zeitschätzungsleistung in einer erwachsenen ADHS-Probe. J. Atten. Disord. doi: 10.1177/1087054716648776. .
PubMed Abstract | CrossRef Full Text | Google Scholar
Price, J. L. (2007). Definition des orbitalen Kortex in Bezug auf spezifische Verbindungen mit limbischen und viszeralen Strukturen und anderen kortikalen Regionen. Annals N.Y. Acad. Sci. 1121, 54-71. doi: 10.1196/annals.1401.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabinak, C. A., MacNamara, A., Kennedy, A. E., Angstadt, M., Stein, M. B., Liberzon, I., et al. (2014). Fokales und abweichendes präfrontales Engagement während der Emotionsregulation bei Veteranen mit posttraumatischer Belastungsstörung. Depress. Anxiety. 31, 851-861. doi: 10.1002/da.22243
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, B., Mehler, B., D’Ambrosio, L. A., and Fried, R. (2010). Die Auswirkungen von Ablenkungen auf junge erwachsene Fahrer mit Aufmerksamkeitsdefizit-Hyperaktivitätsstörung (ADHS). Accid. Anal. Prev. 42, 842-851. doi: 10.1016/j.aap.2009.06.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosch, K. S., Fosco, W. D., Pelham, W. E. Jr., Waxmonsky, J. G., Bubnik, M. G., and Hawk, L. W. Jr. (2016). Verstärkung und Stimulanzien verbessern die mangelnde Reaktionshemmung bei Kindern mit Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung. J. Abnorm. Child Psychol. 44, 309-321. doi: 10.1007/s10802-015-0031-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Lim, L., Ecker, C., Halari, R., Giampietro, V., Simmons, A., et al. (2013). Auswirkungen von Alter und Geschlecht auf neuronale Netzwerke der motorischen Reaktionshemmung: von der Adoleszenz bis zum mittleren Erwachsenenalter. Neuroimage. 83, 690-703. doi: 10.1016/j.neuroimage.2013.06.078
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Oosterlaan, J., Sergeant, J. A., Brandeis, D., and Leeuwen, T. V. (1998). Inhibitorische Dysfunktion bei hyperaktiven Jungen. Behav. Brain Res. 94, 25-32. doi: 10.1016/S0166-4328(97)00166-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Shalom, D. B. (2009). Der mediale präfrontale Kortex und Integration bei Autismus. Neuroscientist. 15, 589-598. doi: 10.1177/1073858409336371
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, J. L., Jamadar, S., Provost, A. L., and Michie, P. T. (2013). Motorische und nicht-motorische Hemmung bei der Go/NoGo-Aufgabe: eine ERP- und fMRI-Studie. Int. J. Psychophysiol. 87, 244-253. doi: 10.1016/j.ijpsycho.2012.07.185
PubMed Abstract | CrossRef Full Text | Google Scholar
Soliman, A. M., and Elfar, R. M. (2017). Falsches Gedächtnis bei Erwachsenen mit ADHS: ein Vergleich zwischen Subtypen und normalen Kontrollen. J. Attent. Disord. 21, 986-996. doi: 10.1177/1087054714556814
PubMed Abstract | CrossRef Full Text | Google Scholar
Sørensen, L., Plessen, K. J., Adolfsdottir, S., and Lundervold, A. J. (2014). Die Spezifität des Stroop-Interferenz-Scores von Fehlern für ADHS bei Jungen. Child Neuropsychol. 20, 677-691. doi: 10.1080/09297049.2013.855716
PubMed Abstract | CrossRef Full Text | Google Scholar
Soto, D., Humphreys, G. W., and Rotshtein, P. (2007). Dissoziierung der neuronalen Mechanismen der gedächtnisbasierten Steuerung der visuellen Auswahl. Proc. Natl. Acad. Sci. U.S.A. 104, 17186-17191. doi: 10.1073/pnas.0703706104
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinelli, S., Joel, S., Nelson, T. E., Vasa, R. A., Pekar, J. J., und Mostofsky, S. H. (2011). Unterschiedliche neuronale Muster sind bei Kindern mit und ohne Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung mit Versuchen verbunden, die hemmenden Fehlern vorausgehen. J. Am. Acad. Child Adolesc. Psychiatry. 50, 705-715. doi: 10.1016/j.jaac.2011.03.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsujimoto, S., Yasumura, A., Yamashita, Y., Torii, M., Kaga, M., and Inagaki, M. (2013). Erhöhte präfrontale Oxygenierung im Zusammenhang mit distraktorresistentem Arbeitsgedächtnis bei Kindern mit Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung (ADHS). Child Psychiatry Hum. Dev. 44, 678-688. doi: 10.1007/s10578-013-0361-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcott, C. M., and Landau, S. (2004). Die Beziehung zwischen Enthemmung und Emotionsregulation bei Jungen mit Aufmerksamkeitsdefizit-Hyperaktivitätsstörung. J. Clin. Child Adolesc. Psychol. 33, 772-782. doi: 10.1207/s15374424jccp3304_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Xiong, K., Zhang, Y., Qiu, M., Zhang, J., Sang, L., Wang, L., et al. (2013). Negative Emotionsregulation bei Patienten mit posttraumatischer Belastungsstörung. PLoS ONE 8:e81957. doi: 10.1371/journal.pone.0081957
PubMed Abstract | CrossRef Full Text | Google Scholar
Yarmolovsky, J., Szwarc, T., Schwartz, M., Tirosh, E., and Geva, R. (2016). Heiße exekutive Kontrolle und Reaktion auf ein Stimulans in einer doppelblinden randomisierten Studie bei Kindern mit ADHS. Eur. Arch. Psychiatry Clin. Neurosci. 267, 73-82. doi: 10.1007/s00406-016-0683-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Yasumura, A., Kokubo, N., Yamamoto, H., Yasumura, Y., Nakagawa, E., Kaga, M., et al. (2014). Neurobehaviorale und hämodynamische Bewertung der Stroop- und umgekehrten Stroop-Interferenz bei Kindern mit Aufmerksamkeitsdefizit-/Hyperaktivitätsstörung. Brain Dev. 36, 97-106. doi: 10.1016/j.braindev.2013.01.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Zsuga, J., Biro, K., Tajti, G., Szilasi, M. E., Papp, C., Juhasz, B., et al. (2016). Proaktive“ Nutzung der Cue-Kontext-Kongruenz für den Aufbau der Belohnungsfunktion des Verstärkungslernens. BMC Neurosci. 17:70. doi: 10.1186/s12868-016-0302-7
PubMed Abstract | CrossRef Full Text | Google Scholar