Eukaryoten haben zwei Hauptarten der Zellteilung: Mitose und Meiose. Die Mitose dient der Bildung neuer Körperzellen für Wachstum und Heilung, während die Meiose der Bildung von Geschlechtszellen (Ei- und Samenzellen) dient. Die Meiose wird in einem späteren Kapitel behandelt.
Der Zellzyklus ist eine geordnete Abfolge von Ereignissen, die das Zellwachstum und die Zellteilung betreffen und bei denen durch Mitose zwei neue Tochterzellen entstehen. Die Dauer des Zellzyklus ist selbst innerhalb der Zellen eines einzelnen Organismus sehr unterschiedlich. Beim Menschen reicht die Häufigkeit des Zellwechsels von wenigen Stunden in der frühen Embryonalentwicklung über durchschnittlich zwei bis fünf Tage bei Epithelzellen bis hin zu einem ganzen Menschenleben ohne Teilung bei spezialisierten Zellen wie kortikalen Neuronen oder Herzmuskelzellen. Auch die Zeit, die eine Zelle in jeder Phase des Zellzyklus verbringt, ist unterschiedlich. Wenn sich schnell teilende Säugetierzellen in Kultur gezüchtet werden (außerhalb des Körpers unter optimalen Wachstumsbedingungen), beträgt die Dauer des Zyklus etwa 24 Stunden. Der zeitliche Ablauf der Ereignisse im Zellzyklus wird durch zellinterne und -externe Mechanismen gesteuert.
Zellen durchlaufen auf dem Weg zur Zellteilung eine Reihe von genau getakteten und sorgfältig regulierten Phasen des Wachstums, der DNA-Replikation und der Teilung, die zwei genetisch identische Zellen hervorbringen. Der Zellzyklus hat zwei Hauptphasen: die Interphase und die mitotische Phase (Abbildung 1). Während der Interphase wächst die Zelle und die DNA wird repliziert. In der mitotischen Phase werden die replizierte DNA und der zytoplasmatische Inhalt getrennt, und die Zelle teilt sich.
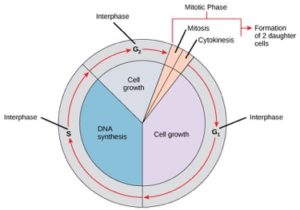
Während der Interphase durchläuft die Zelle normale Prozesse und bereitet sich gleichzeitig auf die Zellteilung vor. Damit eine Zelle von der Interphase in die mitotische Phase übergehen kann, müssen viele interne und externe Bedingungen erfüllt sein. Die drei Stadien der Interphase werden G1, S und G2 genannt.
G1-Phase (erste Lücke)
Das erste Stadium der Interphase wird G1-Phase (erste Lücke) genannt, weil mikroskopisch gesehen nur wenig Veränderung sichtbar ist. Während der G1-Phase ist die Zelle jedoch auf biochemischer Ebene sehr aktiv. Die Zelle akkumuliert die Bausteine der chromosomalen DNA und die dazugehörigen Proteine und sammelt genügend Energiereserven, um die Aufgabe der Replikation jedes Chromosoms im Zellkern zu erfüllen.
S-Phase (DNA-Synthese)
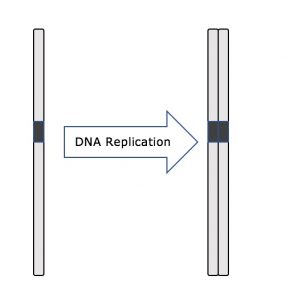
Während der gesamten Interphase bleibt die Kern-DNA in einer halb-kondensierten Chromatin-Konfiguration. In der S-Phase kann die DNA-Replikation durch die Mechanismen fortgesetzt werden, die zur Bildung identischer Paare von DNA-Molekülen – den Schwesternchromatiden – führen, die fest mit der zentromerischen Region verbunden sind (Abbildung 2).
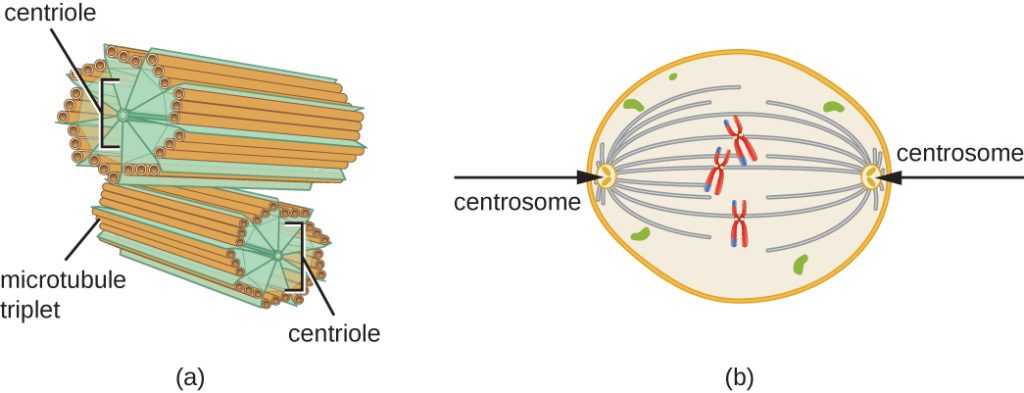
Das Zentrosom wird während der S-Phase ebenfalls dupliziert. Aus den beiden Zentrosomen entsteht die mitotische Spindel, der Apparat, der die Bewegung der Chromosomen während der Mitose koordiniert. Im Zentrum jeder tierischen Zelle sind die Zentrosomen der tierischen Zellen mit einem Paar stäbchenförmiger Objekte, den Zentriolen, verbunden, die im rechten Winkel zueinander stehen. Die Zentriolen helfen bei der Organisation der Zellteilung. In den Zentrosomen anderer eukaryontischer Arten, wie Pflanzen und den meisten Pilzen, sind keine Zentriolen vorhanden.
G2-Phase (Zweite Lücke)
In der G2-Phase füllt die Zelle ihre Energiespeicher auf und synthetisiert Proteine, die für die Chromosomenmanipulation erforderlich sind. Einige Zellorganellen werden dupliziert, und das Zytoskelett wird abgebaut, um Ressourcen für die mitotische Phase bereitzustellen. Während der G2-Phase kann es zu einem zusätzlichen Zellwachstum kommen. Die letzten Vorbereitungen für die mitotische Phase müssen abgeschlossen sein, bevor die Zelle in das erste Stadium der Mitose eintreten kann.
Die mitotische Phase
Um zwei Tochterzellen zu bilden, muss der Inhalt des Zellkerns und des Zytoplasmas geteilt werden. Die mitotische Phase ist ein mehrstufiger Prozess, bei dem die verdoppelten Chromosomen ausgerichtet, getrennt und an die entgegengesetzten Pole der Zelle gebracht werden und die Zelle dann in zwei neue identische Tochterzellen geteilt wird. Der erste Teil der mitotischen Phase, die Mitose, besteht aus fünf Phasen, in denen die Kernteilung durchgeführt wird (Abbildung 5). Der zweite Teil der mitotischen Phase, die so genannte Zytokinese, ist die physische Trennung der zytoplasmatischen Komponenten in zwei Tochterzellen. Obwohl die Stadien der Mitose bei den meisten Eukaryonten ähnlich sind, ist der Prozess der Zytokinese bei Eukaryonten, die Zellwände haben, wie z. B. Pflanzenzellen, ganz anders.
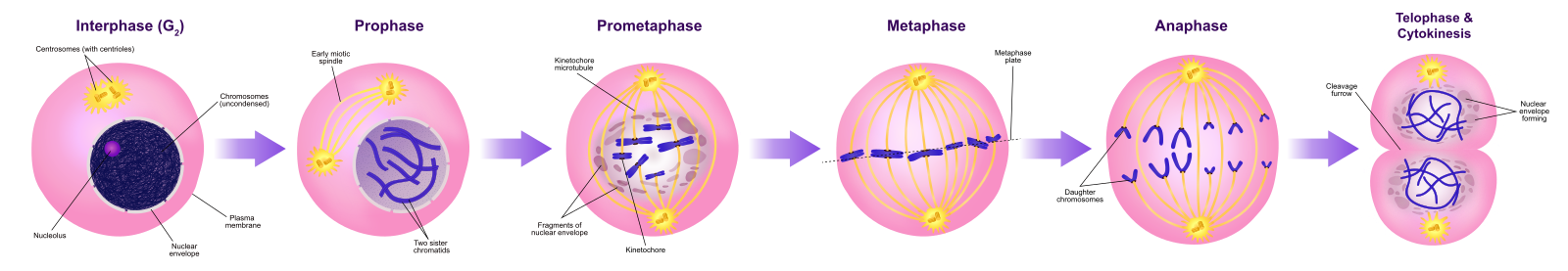
Prophase
Während der Prophase, der „ersten Phase“, beginnt die Kernhülle in kleine Bläschen zu zerfallen, und die membranösen Organellen (wie der Golgi-Apparat und das endoplasmatische Retikulum) fragmentieren und verteilen sich zu den Rändern der Zelle. Der Nukleolus verschwindet. Die Zentrosomen beginnen, sich zu den gegenüberliegenden Polen der Zelle zu bewegen. Mikrotubuli, die die mitotische Spindel bilden werden, erstrecken sich zwischen den Zentrosomen und schieben sie weiter auseinander, während sich die Mikrotubuli-Fasern verlängern. Die Schwesterchromatiden beginnen sich mit Hilfe von Kondensinproteinen enger zu wickeln und werden unter dem Lichtmikroskop sichtbar.
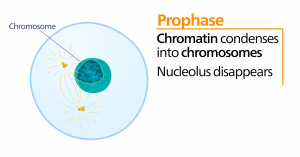
Prometaphase
Während der Prometaphase, der „ersten Veränderungsphase“, schreiten viele Prozesse, die in der Prophase begonnen wurden, weiter voran. Die Reste der Kernhülle zersplittern. Die mitotische Spindel entwickelt sich weiter, da sich mehr Mikrotubuli zusammenfinden und sich über die Länge des ehemaligen Kernbereichs erstrecken. Die Chromosomen werden kondensierter und diskreter. Jedes Schwesterchromatid entwickelt eine Proteinstruktur, die Kinetochore genannt wird, in der zentromerischen Region.
Die Proteine des Kinetochors ziehen Mikrotubuli der mitotischen Spindel an und binden sie. Wenn sich die Spindelmikrotubuli von den Zentrosomen ausbreiten, kommen einige dieser Mikrotubuli mit den Kinetochoren in Kontakt und binden sich fest daran. Sobald eine mitotische Faser an einem Chromosom anhaftet, wird das Chromosom so ausgerichtet, dass die Kinetochoren der Schwesterchromatiden den gegenüberliegenden Polen zugewandt sind. Schließlich werden alle Schwesterchromatiden über ihre Kinetochoren an Mikrotubuli von gegenüberliegenden Polen befestigt. Spindelmikrotubuli, die nicht an den Chromosomen angreifen, werden als polare Mikrotubuli bezeichnet. Diese Mikrotubuli überlagern sich in der Mitte zwischen den beiden Polen und tragen zur Zelldehnung bei. Astrale Mikrotubuli befinden sich in der Nähe der Pole, helfen bei der Spindelausrichtung und sind für die Regulierung der Mitose erforderlich.

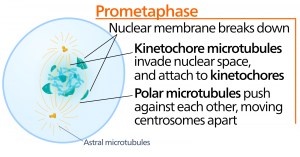
Metaphase
In der Metaphase, der „Veränderungsphase“, sind alle Chromosomen in einer Ebene ausgerichtet, die als Metaphasenplatte oder Äquatorialebene bezeichnet wird und in der Mitte zwischen den beiden Polen der Zelle liegt. Die Schwesterchromatiden sind noch immer durch Kohäsinproteine fest miteinander verbunden. Zu diesem Zeitpunkt sind die Chromosomen maximal kondensiert.
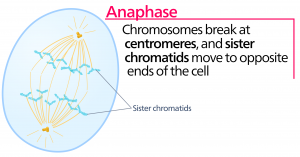
Anaphase
In der Anaphase, der „Aufwärtsphase“, werden die Kohäsinproteine abgebaut, und die Schwesterchromatiden trennen sich am Zentromer. Jedes Chromatid, das nun als Chromosom bezeichnet wird, wird schnell zum Zentrosom gezogen, an dem seine Mikrotubuli befestigt sind. Die Zelle wird sichtbar länglich (oval), da die polaren Mikrotubuli an der Metaphaseplatte, wo sie sich überlappen, gegeneinander gleiten.
Telophase
Während der Telophase, der „Abstandsphase“, erreichen die Chromosomen die gegenüberliegenden Pole und beginnen zu dekondensieren (sich zu entwirren), indem sie sich in eine Chromatinkonfiguration entspannen. Die mitotischen Spindeln werden in Tubulinmonomere depolymerisiert, die zum Aufbau der Zytoskelettkomponenten für jede Tochterzelle verwendet werden. Kernhüllen bilden sich um die Chromosomen, und Nukleosomen erscheinen im Kernbereich.
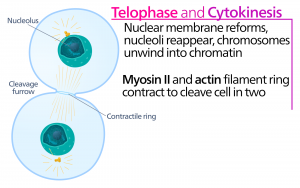
Zytokinese
Die Zytokinese oder „Zellbewegung“ ist das zweite Hauptstadium der mitotischen Phase, in dem die Zellteilung durch die physische Trennung der zytoplasmatischen Komponenten in zwei Tochterzellen abgeschlossen wird. Die Teilung ist erst dann abgeschlossen, wenn sich die Zellbestandteile geteilt und vollständig in die beiden Tochterzellen getrennt haben. Obwohl die Stadien der Mitose bei den meisten Eukaryonten ähnlich sind, ist der Prozess der Zytokinese bei Eukaryonten, die Zellwände haben, wie z. B. Pflanzenzellen, ganz anders.
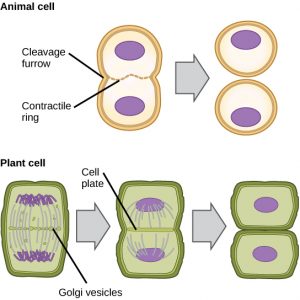
In Zellen wie tierischen Zellen, die keine Zellwände haben, folgt die Zytokinese auf den Beginn der Anaphase. Ein kontraktiler Ring aus Aktinfilamenten bildet sich direkt innerhalb der Plasmamembran an der ehemaligen Metaphasenplatte (Abbildung 12). Die Aktinfilamente ziehen den Äquator der Zelle nach innen und bilden einen Spalt. Dieser Spalt oder „Riss“ wird als Spaltfurche bezeichnet. Die Furche vertieft sich, wenn sich der Aktinring zusammenzieht, und schließlich wird die Membran in zwei Teile gespalten.
In Pflanzenzellen muss sich eine neue Zellwand zwischen den Tochterzellen bilden. Während der Interphase sammelt der Golgi-Apparat Enzyme, Strukturproteine und Glukosemoleküle an, bevor er in Bläschen zerfällt und sich in der sich teilenden Zelle verteilt (Abbildung 12). Während der Telophase werden diese Golgi-Bläschen auf Mikrotubuli transportiert, um an der Metaphasenplatte einen Phragmoplast (eine Bläschenstruktur) zu bilden. Dort verschmelzen die Vesikel und wachsen vom Zentrum aus in Richtung der Zellwände zusammen; diese Struktur wird als Zellplatte bezeichnet. Wenn weitere Bläschen verschmelzen, vergrößert sich die Zellplatte, bis sie an der Peripherie der Zelle mit den Zellwänden verschmilzt. Enzyme verwenden die Glukose, die sich zwischen den Membranschichten angesammelt hat, um eine neue Zellwand aufzubauen. Die Golgi-Membranen werden zu Teilen der Plasmamembran auf beiden Seiten der neuen Zellwand.
Zusammenfassung von Mitose und Zytokinese
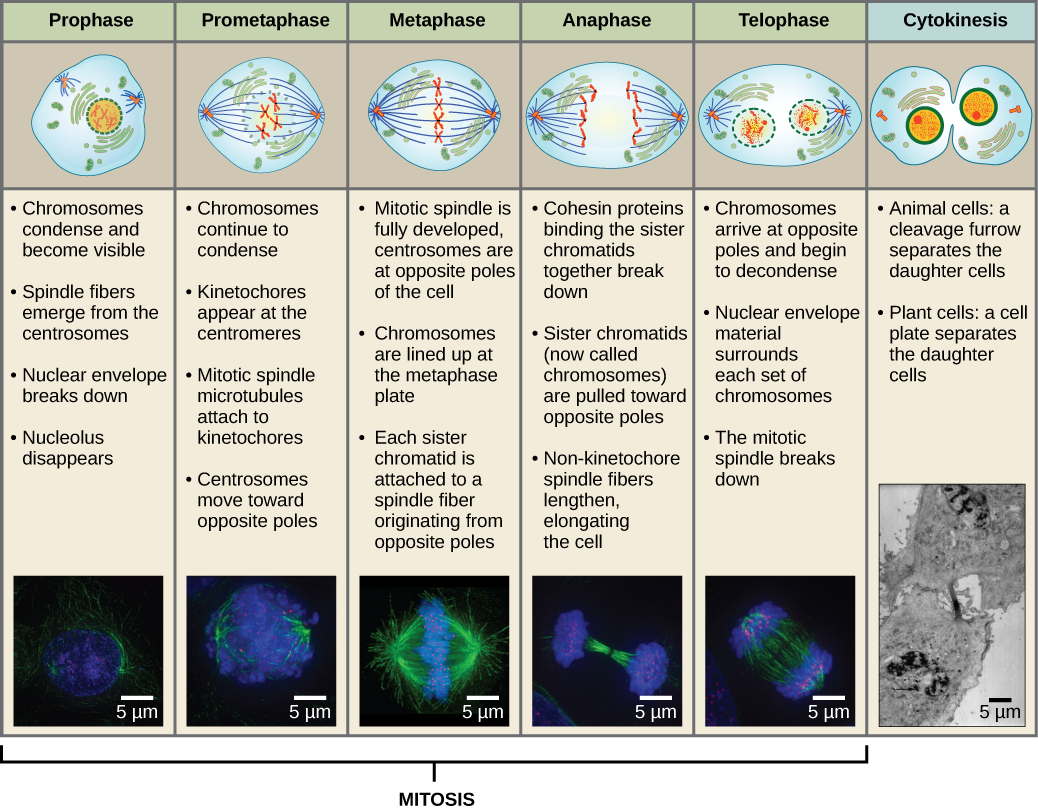
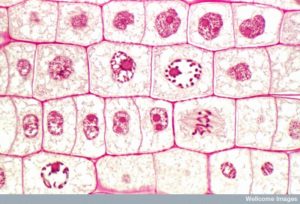
Abbildung 13 Die Mitose ist in fünf Stadien unterteilt – Prophase, Metaphase, Anaphase und Telophase. Die Bilder unten wurden mit Hilfe der Fluoreszenzmikroskopie von Zellen aufgenommen, die künstlich mit Fluoreszenzfarbstoffen angefärbt wurden: blaue Fluoreszenz zeigt die DNA (Chromosomen) und grüne Fluoreszenz die Mikrotubuli (Spindelapparat) an. (Credit „Mitose-Zeichnungen“: Modifikation der Arbeit von Mariana Ruiz Villareal; Credit „Mikrografien“: Modifikation der Arbeit von Roy van Heesbeen; Credit „Zytokinese-Mikrografie“: Wadsworth Center/New York State Department of Health; scale-bar data from Matt Russell)
G0 Phase
Nicht alle Zellen halten sich an das klassische Zellzyklusmuster, bei dem eine neu gebildete Tochterzelle sofort in die Interphase eintritt, dicht gefolgt von der mitotischen Phase. Zellen, die sich in der G0-Phase befinden, bereiten sich nicht aktiv auf eine Teilung vor. Die Zelle befindet sich in einem ruhenden (inaktiven) Stadium, nachdem sie den Zellzyklus verlassen hat. Einige Zellen befinden sich vorübergehend in der G0-Phase, bis ein externes Signal den Beginn der G1-Phase auslöst. Andere Zellen, die sich nie oder nur selten teilen, wie z. B. reife Herzmuskel- und Nervenzellen, verbleiben dauerhaft in G0.