Introducción
Un trabajo anterior de nuestro laboratorio (Shalom, 2009) argumentaba que el procesamiento neurocognitivo puede dividirse en tres niveles: (1) un nivel básico que implica el procesamiento cognitivo, emocional y sensoriomotor primario. Por ejemplo, un sonido fuerte e inesperado que se percibe en los sistemas sensoriales primarios puede desencadenar respuestas emocionales fisiológicas, como el miedo (latidos rápidos del corazón, etc.). (2) un nivel integrador, que combina la salida de todos los procesos primarios del nivel básico y forma un significado, una experiencia o un comportamiento global y coherente. Por ejemplo, las representaciones mentales de los distintos elementos primarios que constituyen la respuesta emocional del miedo dan como resultado una sensación consciente de tener miedo. (3) Un nivel lógico, que forma reglas lógicas abstractas (reglas «si-entonces») a partir del nivel básico (por ejemplo, «Si tengo latidos rápidos del corazón y sudor frío, entonces podría tener miedo»), y también está involucrado en la selección e inhibición (por ejemplo, el control de los impulsos emocionales). Esta arquitectura de tres niveles se aplicó a cuatro dominios psicológicos generales: emoción, memoria, sensación-percepción y motor. Shalom (2009) se centró en el segundo nivel, el integrador, y su relación con los cuatro dominios psicológicos. También argumentó que estos cuatro tipos de integración están subservidos por cuatro subáreas diferentes de la corteza prefrontal medial: Área de Brodmann (BA) 11 (percepción), BA 10 (memoria), BA 9 (emoción) y BA 8 (motor). Por último, presentó pruebas de que un déficit selectivo en estas áreas BA y estos tipos de procesos integradores subyacen a algunos de los déficits comunes en los TEA (trastornos del espectro autista).
La presente revisión intenta realizar un análisis similar para el córtex prefrontal lateral (LPFC), pasando de las áreas prefrontales mediales relevantes a sus homólogas laterales: BA lateral 11/BA 47 (percepción), BA lateral 10/BA 46 (memoria), BA lateral 9 (emoción) y BA lateral 8 (motor) (Figura 1), (cf. la distinción entre los clusters 1, 2, 3 y 4 del cíngulo anterior en Beckmann et al. (2009). Aporta pruebas de que una atipicidad selectiva en estas áreas del BA y un déficit selectivo en los procesos de selección/inhibición en estos cuatro dominios cognitivos están implicados en algunos de los déficits comunes en el TDAH (trastorno por déficit de atención con hiperactividad), como las dificultades para inhibir las respuestas motoras (p. ej, incapacidad para inhibir movimientos inapropiados), la concentración perceptiva (por ejemplo, incapacidad para ignorar la distracción) y las reacciones emocionales (por ejemplo, incapacidad para controlar los impulsos) (Barkley, 1997; Nigg, 1999).
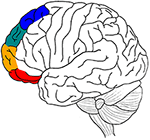
Figura 1. Resumen gráfico de las principales hipótesis anatómicas del artículo. Procesamiento motor en azul, emoción en verde, memoria en naranja y sensorial-perceptivo en rojo.
Percepción
Selección e inhibición perceptiva y BA 11/47 lateral
Hay evidencias de que la selección e inhibición perceptiva se apoya en redes neuronales que implican la parte lateral de BA 11/47 en el LPFC. En un modelo propuesto por Zsuga et al. (2016), se sugiere que la corteza orbitofrontal (OFC) desempeña un papel importante en la selección de estímulos visuales de acuerdo con los requisitos de la tarea.En concreto, sugieren que la parte medial de la OFC desempeña un papel central en la integración y categorización de la información sobre los estímulos del entorno y su contexto, mientras que la parte lateral de la OFC está involucrada en la asignación y actualización de los parámetros de selección de acuerdo con los valores específicos de la tarea de los estímulos disponibles. En apoyo de esta afirmación, en un estudio, Howard y Kahnt (2017) demostraron que la parte lateral del BA 11 está implicada en la codificación de valores dirigidos a la meta de los estímulos olfativos. Presentaron a participantes hambrientos dos olores de comida preferidos y les pidieron que eligieran si querían oler uno u otro. Más tarde, los participantes recibieron uno de los dos alimentos para comer y se les permitió elegir entre los dos olores una vez más. El cambio en la preferencia hacia el olor no saturado se reflejó en un cambio de actividad en la parte lateral de la BA 11/47. Además, se ha demostrado que la BA 11/47 lateral está implicada en el rechazo de estímulos irrelevantes (Kaufman et al., 2016).
También se ha demostrado que la BA 11/47 lateral está implicada en el mantenimiento de los estímulos visuales en la memoria de trabajo y la orientación de la selección visual. En un estudio, Soto et al. (2007), compararon dos condiciones de una selección visual cebada en la que el cebador se mostraba una o dos veces. La activación lateral del BA 11/47 fue mayor cuando el cebador se mostró dos veces, sin embargo, se encontró una reducción de la activación cuando estímulos similares se repitieron pasivamente dos veces (sin una fase de selección visual, y por lo tanto sin relevancia para la tarea).
Por último, según Price (2007), la BA 11 lateral forma parte de la red orbital sensorial.
El TDAH y la selección e inhibición visual
Hay pruebas de que los niños y adultos con TDAH tienen problemas para inhibir estímulos visuales y auditivos irrelevantes. Por ejemplo, los adultos con TDAH han mostrado más efectos de interferencia (es decir, problemas para inhibir los estímulos irrelevantes) que los controles cuando se les pide que completen una tarea de recuerdo mientras ignoran un ruido de fondo (Pelletier et al., 2016). Además, se encontró una correlación positiva entre el rendimiento de la tarea en condiciones de sonido irrelevante y el alcance de los síntomas atencionales reportados por los pacientes en una escala de síntomas clínicos. En otro estudio, se pidió a adultos con TDAH y a adultos de control que mantuvieran una conversación telefónica mientras conducían. El grupo con TDAH mostró habilidades de conducción significativamente más pobres durante la condición de conversación telefónica que en la condición de silencio en comparación con los controles (Reimer et al., 2010). Otros estudios han mostrado una pobre inhibición visual en individuos con TDAH. Por ejemplo, Forster et al. (2014) y Forster y Lavie (2016) descubrieron que los adultos con TDAH realizaban una tarea de búsqueda de letras más lentamente cuando había personajes de dibujos animados irrelevantes pero salientes en la pantalla, en mayor medida que los controles. De manera relacionada, la tarea de interferencia de Stroop requiere que los participantes atiendan una dimensión específica de un estímulo visual mientras ignoran otra. Los estudios encontraron que los niños con TDAH tienen un desempeño más pobre en la condición de interferencia Stroop que los controles (Sørensen et al., 2014), y que su desempeño se correlacionó con sus síntomas de inatención e hiperactividad (Ikeda et al., 2013).
TDAH, selección e inhibición visual y BA 11/47 lateral
Además, varios estudios encontraron evidencia específica de activación atípica en BA 11/47 lateral en individuos con TDAH que realizan tareas de selección/inhibición visual. Tsujimoto et al. (2013) pidieron a niños con y sin TDAH que realizaran una tarea de memoria de trabajo con y sin distracción. Los participantes con TDAH mostraron un rendimiento conductual significativamente más pobre, en particular bajo distracción, y mostraron niveles significativamente más altos de activación en el BA 11/47 lateral que los controles. En un estudio diferente, Yasumura et al. (2014) evaluaron a niños con TDAH y a niños de control en condiciones de interferencia Stroop y de interferencia Stroop inversa, y encontraron que en ambas condiciones los niños con TDAH tuvieron un desempeño más pobre y que el desempeño se correlacionó con la actividad lateral de BA 47. En otro estudio en el que se presentó un distractor saliente pero irrelevante durante una tarea de estimación del tiempo, se encontró que mientras el distractor ayudaba a los participantes con TDAH en el mismo grado que a los controles, la activación lateral de BA 11 que acompañaba a la aparición del distractor era significativamente mayor (Pretus et al, 2016).
Memoria
Selección e inhibición de la memoria y BA 10/46 lateral
Si bien no existe un consenso generalizado de que BA10/46 participe en las funciones de la memoria episódica, hay muchas pruebas que apoyan un papel de BA 10/46 en la selección e inhibición de la información de la memoria. Por ejemplo, Kim (2011), analizó 74 estudios que comparaban el recuerdo y el olvido (es decir, los elementos recordados con éxito frente a los distractores juzgados erróneamente como aparecidos), y encontró que la mayor parte de los estudios se superponían en la parte frontal de BA 46, cerca de su unión con BA 10 lateral. En un estudio posterior, Kim (2013) realizó un meta-análisis de 70 estudios centrados en elementos nuevos y antiguos (es decir, aciertos frente a rechazos correctos). Los resultados muestran que las comparaciones nuevas>viejas producían activaciones que se extendían por la parte media del prefrontal lateral izquierdo. Sin embargo, al centrarse en los estudios de memoria de origen en los que se exige a los participantes que recuperen no sólo el ítem objetivo sino también un detalle adicional relacionado con el ítem de la fase de aprendizaje (es decir, cuando las demandas de selección son mayores), se observó una activación específica en la parte frontal de la BA46, incluida la parte rostral de la BA10.
Una línea de evidencia diferente proviene de un meta-análisis realizado por Gilbert et al. (2006), en el que se analizaron 104 estudios centrados en la BA 10, y se encontró que la mayoría de los estudios que implicaban la parte lateral de la BA 10 evaluaban la memoria de trabajo y la recuperación de la memoria episódica, mientras que los estudios que implicaban la parte medial de la BA 10 evaluaban la mentalización (es decir, atender a las propias emociones y estados mentales o a los de otros agentes).
Además, Cohen et al. (2014), emplearon la tarea de recuerdo dirigido por el valor, en la que se presenta a los participantes listas de palabras en las que se asigna a algunas palabras más valor que a otras, lo que anima a los participantes a intentar y seleccionar elementos de mayor valor sobre los de menor valor durante la codificación. Los resultados muestran una activación específica de la BA46/10 lateral derecha, y una activación muy pronunciada de la BA46/10 lateral izquierda, cuando aparecía una pista que predecía una palabra de alto valor.
El TDAH y la selección e inhibición de la memoria
Hay varios estudios que han encontrado evidencia de dificultades en la selección e inhibición de la información de la memoria en el TDAH. Por ejemplo, en un estudio de Pollak et al. (2007), adultos con y sin TDAH realizaron una difícil tarea de aprendizaje de listas en la que tenían que memorizar y recordar cinco listas de palabras en ocho ocasiones diferentes. Los resultados muestran que los participantes con TDAH cometieron más recuerdos dobles y errores de intrusión (recordar palabras de la lista equivocada), que son ejemplos de una dificultad para inhibir la recuperación de elementos irrelevantes para la tarea. En un estudio similar, Soliman y Elfar (2017), pidieron a adultos con y sin TDAH que aprendieran ocho listas y luego se les presentó una tarea de reconocimiento. El grupo con TDAH reconoció menos palabras correctamente, produjo más respuestas positivas falsas (es decir, seleccionó erróneamente elementos que no fueron aprendidos) y se mostró más confiado en sus errores. En un estudio diferente, Castel et al. (2011), pidieron a niños con y sin TDAH que realizaran el paradigma de recuerdo dirigido por valores mencionado anteriormente. Los resultados muestran que los niños con TDAH recordaron tantos ítems como los controles, sin embargo fueron menos capaces de seleccionar qué ítems recordar.
TDAH, Selección e Inhibición de la Memoria y BA Lateral 10/46
La búsqueda bibliográfica sólo arrojó un estudio de neuroimagen que evaluaba la selección o inhibición de las funciones de memoria en el TDAH. En un estudio de Depue et al. (2010), se enseñó a adultos con y sin TDAH una relación entre pares de caras e imágenes hasta su dominio. Posteriormente se les mostró una imagen de una cara y se les pidió que pensaran en la imagen relacionada o que evitaran que la imagen relacionada llegara a la conciencia. Los resultados muestran que los participantes con TDAH activaron el BA 10 lateral menos que los controles al comparar la condición de «no pensar» con la de «pensar».
Emoción
Selección e inhibición de emociones y BA 9 lateral
Hay muchas pruebas que apoyan la relación entre la regulación de las emociones (la inhibición de ciertas emociones y la selección de otras) y el BA 9 lateral. Por ejemplo, en un estudio realizado por Hallam et al. (2014), los participantes vieron películas que inducían emociones y se les pidió que reprimieran o reapreciaran su reacción emocional, o que simplemente vieran la película. Los resultados mostraron que tanto la reapreciación como la supresión, en comparación con la condición pasiva, mostraban una activación de la BA 9 lateral. De forma similar, dos estudios compararon la reapreciación y la observación pasiva de imágenes en personas con TEPT y controles. Ambos estudios encontraron que la BA 9 lateral se activaba más durante la condición de reappraisal, pero en menor medida en las personas con TEPT (Xiong et al., 2013; Rabinak et al., 2014).
Además, se utilizaron dos meta-análisis diferentes para examinar los estudios de fMRI que evaluaban la regulación de la emoción. Buhle et al. (2014) encontraron que la revaloración que implicaba una regulación descendente del afecto negativo activaba consistentemente la BA 9 lateral; Frank et al. (2014) encontraron que dicha revaloración iba acompañada de una mayor activación en la BA 9 lateral y una menor activación en la amígdala.
El TDAH y la selección e inhibición de emociones
Varios estudios encontraron déficits de regulación de las emociones en niños con TDAH. Por ejemplo, varios estudios muestran que a los niños con TDAH les cuesta más reprimir sus emociones que a sus compañeros de desarrollo típico (DT). Por ejemplo, en un estudio los niños se sometieron a una tarea de competencia entre pares, en la que a la mitad se le pidió que ocultara sus emociones y a la otra mitad no. Según las evaluaciones realizadas por jueces ingenuos, los resultados muestran que los niños con TDAH fueron menos capaces de ocultar sus emociones que sus compañeros (Walcott y Landau, 2004). Además, un estudio que evaluó el rendimiento de los niños con TDAH en una tarea Stroop emocional, encontró que los niños con TDAH tenían más dificultades para inhibir las respuestas a las caras enojadas y frustradas (Yarmolovsky et al., 2016).
Además, en un meta-análisis de la regulación de las emociones en el TDAH realizado por Graziano y García (2016), encontraron cuatro dominios de interés, a saber: reconocimiento (es decir, capacidad de procesar e inferir las emociones de los demás, así como las propias), la reactividad (es decir, el umbral, la intensidad y la duración de la propia excitación afectiva), la regulación (es decir, responder eficazmente a la reactividad emocional de una manera flexible que facilite el funcionamiento adaptativo) y la empatía (es decir, la capacidad de experimentar el estado afectivo de otro y/o expresar preocupación por la posición de otro). Los resultados muestran que, aunque los cuatro dominios parecen ser atípicos en la población con TDAH, las deficiencias más notables se dan en los dominios de reacción y regulación. Esto significa que (especialmente) los niños con TDAH tienden a reaccionar más rápidamente, más intensamente y durante más tiempo a las situaciones aversivas, y son menos capaces de regular estas emociones incluso cuando intentan hacerlo.
El TDAH, la selección de emociones y la inhibición y el BA 9 lateral
Hay varios estudios que muestran atipicidades específicas en la activación del BA 9 lateral en participantes con TDAH que realizan tareas que requieren regulación de emociones. Por ejemplo, en un estudio realizado por Passarotti et al. (2010a), niños con trastorno bipolar, niños con TDAH y un grupo de controles de la misma edad realizaron una tarea Stroop de valencia emocional. En esta tarea, se presentaron palabras positivas, neutras y negativas, y los participantes debían responder según el color de la palabra mientras ignoraban su significado. Los resultados muestran que el grupo bipolar tenía dificultades para ignorar las palabras positivas, mientras que el grupo con TDAH tenía dificultades para ignorar las palabras negativas. Curiosamente, la activación lateral del BA 9 fue más pronunciada para la condición de la tarea que se realizó con éxito, es decir, para las palabras negativas en el grupo bipolar, y para las palabras positivas en el grupo TDAH. Además, en una tarea emocional basada en Stroop, Hwang et al. (2015), pidieron a niños con y sin TDAH que realizaran una tarea Stroop numérica después de que se les mostraran imágenes que eran positivas, neutras o negativas. Aunque los datos conductuales no mostraron diferencias significativas entre las diferentes condiciones emocionales, sí encontraron que los niños con TDAH eran menos capaces de reclutar el BA 9 lateral cuando realizaban la tarea, un rasgo que también se correlacionó con la gravedad de los síntomas, tal y como se midió con la escala de padres de Conner.
Motor
Selección e inhibición motora y BA 8 lateral
Hay muchas evidencias de que la inhibición de las acciones motoras se apoya en las redes neuronales del BA 8 lateral. La medida más común de la inhibición motora es la Tarea de la Señal de Parada (SST) en la que se pide a los participantes que pulsen una tecla cuando se presenta un estímulo, pero que se abstengan de ejecutar esa pulsación de la tecla si el estímulo es seguido en breve por una señal. Los estudios demuestran que la inhibición de la pulsación de teclas implica la activación de la BA 8 lateral (Matthews et al., 2005; Smith et al., 2013; Hughes et al., 2014). También se ha demostrado que la BA 8 lateral se activa específicamente cuando la selección de una respuesta motora requería atención o implicaba un conflicto. Por ejemplo, Enríquez-Geppert et al. (2013), pidieron a los participantes que pulsaran una tecla cuando aparecía un estímulo en la pantalla, sin embargo, la tecla de respuesta cambió en algunos de los ensayos. Los experimentadores encontraron que el BA 8 lateral se activaba específicamente en los ensayos en los que la tecla de respuesta era diferente. Del mismo modo, Lenartowicz et al. (2011), utilizaron una tarea de ir/no ir en la que después de algunas sesiones de práctica, los estímulos cambiaron de manera que la señal anterior de ir se convirtió en una señal de no ir (requiriendo más inhibición). Encontraron que la BA 8 lateral se activaba específicamente en esos ensayos. En un estudio similar, Albares et al. (2014), utilizaron una versión de una tarea de ir/no ir en la que se presentaba un prime antes de los estímulos de ir y no ir. Sin embargo, mientras que una luz verde indicaba que seguiría un estímulo go, una luz roja significaba que podía darse cualquiera de las tres condiciones, un estímulo go, un estímulo no-go o ningún estímulo. Los investigadores razonaron que la condición de luz roja implicaba una planificación motora más compleja que la luz verde, y descubrieron que el BA 8 lateral era, de hecho, más activo durante esa condición.
El TDAH y la selección e inhibición motora
Hay pruebas sólidas de déficits de selección e inhibición motora en niños con TDAH. Muchos estudios hasta la fecha han demostrado que los niños, adolescentes y adultos con TDAH muestran un rendimiento más pobre en diferentes versiones de la tarea de señal de parada (por ejemplo, Rubia et al., 1998; Lee et al., 2016; Bialystok et al., 2017; Dekkers et al., 2017), y que cuando los niños con TDAH son adecuadamente medicados y motivados utilizando un refuerzo eficaz, estas diferencias pueden desaparecer (Rosch et al., 2016). Además, en un estudio muy amplio realizado por Crosbie et al. (2013), se midieron los síntomas del TDAH en más de 16.000 niños de la población general que realizaron el SST. Los resultados mostraron que los individuos con mayores puntuaciones de rasgos de TDAH tenían una peor inhibición de la respuesta, una latencia de respuesta más lenta y una mayor variabilidad, y también que esta tendencia era altamente heredable. De manera similar, Alderson et al. (2007), realizaron un meta-análisis centrado en los niños con TDAH y el SST, y hallaron mayores tiempos medios de respuesta, mayor variabilidad de respuesta y mayor tiempo de respuesta de la señal de parada en el grupo con TDAH.
Toma de conciencia del TDAH, selección e inhibición motora y BA 8 lateral
También hay evidencia considerable que muestra atípicos específicos en la activación del BA 8 lateral en participantes con TDAH que realizan tareas que requieren selección e inhibición motora. Por ejemplo, dos meta-análisis separados han explorado estudios de RMf que evalúan la inhibición motora en niños y adultos con TDAH (Hart et al., 2013; Rubia et al., 2013). Ambos meta-análisis encontraron que una menor activación en el BA 8 lateral estaba relacionada con las medidas de inhibición motora, y el estudio de Rubia et al. (2013), también encontró que el metilfenidato aumentaba la activación en el BA 8 lateral, lo que estaba relacionado con un mejor rendimiento en las tareas motoras. Además, los estudios que emplearon diferentes versiones del SST y de la tarea de ir/no ir encontraron una menor activación en el BA 8 lateral en el grupo de TDAH durante una condición simple de no ir (Passarotti et al., 2010b; Mulligan et al., 2011), momentos antes de que se produjera un error (Spinelli et al., 2011), así como durante una condición de libre elección en la que los participantes elegían si pulsar o no un botón cuando se presentaba un estímulo (Karch et al., 2010).
Resumen
El presente trabajo presenta evidencias de que la parte lateral de la corteza prefrontal está implicada en la inhibición, selección y manipulación de la información motora, emocional, de memoria y perceptivo-sensorial, una función que resulta útil en la vida cotidiana por la capacidad de ignorar distracciones, seleccionar y recuperar información específica de nuestra memoria, identificar y regular nuestro estado emocional o planificar una respuesta motora adecuada a la situación. Utiliza este modelo para clasificar los desafíos que encuentran las personas con TDAH, vinculando diferentes tipos de dificultades en la selección/inhibición a diferentes subáreas de la corteza prefrontal lateral.
Contribuciones del autor
El autor confirma ser el único colaborador de este trabajo y lo aprobó para su publicación.
Declaración de conflicto de intereses
El autor declara que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un potencial conflicto de intereses.
Albares, M., Lio, G., Criaud, M., Anton, J. L., Desmurget, M., y Boulinguez, P. (2014). La corteza frontal medial dorsal media la inhibición motora automática en contextos inciertos: evidencia de estudios combinados de fMRI y EEG. Hum. Brain Mapp. 35, 5517-5531. doi: 10.1002/hbm.22567
PubMed Abstract | CrossRef Full Text | Google Scholar
Alderson, R. M., Rapport, M. D., y Kofler, M. J. (2007). El trastorno por déficit de atención/hiperactividad y la inhibición conductual: una revisión meta-analítica del paradigma de la señal de parada. J. Abnorm. Child Psychol. 35, 745-758. doi: 10.1007/s10802-007-9131-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Barkley, R. A. (1997). Inhibición conductual, atención sostenida y funciones ejecutivas: construcción de una teoría unificadora del TDAH. Psychol. Bull. 121:65. doi: 10.1037/0033-2909.121.1.65
PubMed Abstract | CrossRef Full Text | Google Scholar
Beckmann, M., Johansen-Berg, H., y Rushworth, M. F. (2009). Connectivity-based parcellation of human cingulate cortex and its relation to functional specialization. J. Neurosci. 29, 1175-1190. doi: 10.1523/JNEUROSCI.3328-08.2009
PubMed Abstract | CrossRef Full Text | Google Scholar
Bialystok, E., Hawrylewicz, K., Wiseheart, M., y Toplak, M. (2017). Interacción del bilingüismo y el trastorno por déficit de atención/hiperactividad en adultos jóvenes. Bilingualism 20, 588-601. doi: 10.1017/S1366728915000887
PubMed Abstract | CrossRef Full Text | Google Scholar
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., et al. (2014). Revalorización cognitiva de la emoción: un meta-análisis de estudios de neuroimagen en humanos. Cereb. Cortex 24, 2981-2990. doi: 10.1093/cercor/bht154
PubMed Abstract | CrossRef Full Text | Google Scholar
Castel, A. D., Lee, S. S., Humphreys, K. L., and Moore, A. N. (2011). Capacidad de memoria, control selectivo y recuerdo dirigido por valores en niños con y sin trastorno por déficit de atención/hiperactividad (TDAH). Neuropsicología 25:15. doi: 10.1037/a0020298
PubMed Abstract | CrossRef Full Text | Google Scholar
Cohen, M. S., Rissman, J., Suthana, N. A., Castel, A. D., y Knowlton, B. J. (2014). La modulación basada en el valor de la codificación de la memoria implica el compromiso estratégico de las regiones de procesamiento semántico fronto-temporal. Cogn. Affect. Behav. Neurosci. 14, 578-592. doi: 10.3758/s13415-014-0275-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crosbie, J., Arnold, P., Paterson, A., Swanson, J., Dupuis, A., Li, X., et al. (2013). Inhibición de respuesta y rasgos de TDAH: correlatos y heredabilidad en una muestra comunitaria. J. Abnorm. Child Psychol. 41, 497-507. doi: 10.1007/s10802-012-9693-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Dekkers, T. J., van Rentergem, J. A. A., Koole, A., van den Wildenberg, W. P., Popma, A., Bexkens, A., et al. (2017). Efectos del tiempo en la tarea en niños con y sin TDAH: ¿agotamiento de los recursos ejecutivos o agotamiento de la motivación? Eur. Child Adolesc. Psychiatry. 26, 1-11. doi: 10.1007/s00787-017-1006-y
CrossRef Full Text | Google Scholar
Depue, B. E., Burgess, G. C., Willcutt, E. G., Ruzic, L., y Banich, M. T. (2010). Control inhibitorio de la recuperación de la memoria y el procesamiento motor asociado con la corteza prefrontal lateral derecha: evidencia de los déficits en individuos con TDAH. Neuropsychologia. 48, 3909-3917. doi: 10.1016/j.neuropsychologia.2010.09.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Enríquez-Geppert, S., Eichele, T., Specht, K., Kugel, H., Pantev, C., y Huster, R. J. (2013). Parcelación funcional de las cortezas frontal inferior y midcingulada en un paradigma de cambio de flanco-parada. Hum. Brain Mapp. 34, 1501-1514. doi: 10.1002/hbm.22002
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., and Lavie, N. (2016). Estableciendo el rasgo de distracción de la atención. Psychol. Sci. 27, 203-212. doi: 10.1177/0956797615617761
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., Robertson, D. J., Jennings, A., Asherson, P., y Lavie, N. (2014). Tapando el déficit de atención: la carga perceptiva contrarresta el aumento de la distracción en el TDAH. Neuropsicología 28:91. doi: 10.1037/neu0000020
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank, D. W., Dewitt, M., Hudgens-Haney, M., Schaeffer, D. J., Ball, B. H., Schwarz, N. F., et al. (2014). Regulación de la emoción: meta-análisis cuantitativo de la activación y desactivación funcional. Neurosci. Biobehav. Rev. 45, 202-211. doi: 10.1016/j.neubiorev.2014.06.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Gilbert, S. J., Spengler, S., Simons, J. S., Steele, J. D., Lawrie, S. M., Frith, C. D., et al. (2006). Functional specialization within rostral prefrontal cortex (area 10): a meta-analysis. J. Cogn. Neurosci. 18, 932-948. doi: 10.1162/jocn.2006.18.6.932
PubMed Abstract | CrossRef Full Text | Google Scholar
Graziano, P. A., y García, A. (2016). Trastorno por déficit de atención e hiperactividad y desregulación emocional infantil: un meta-análisis. Clin. Psychol. Rev. 46, 106-123. doi: 10.1016/j.cpr.2016.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Hallam, G. P., Webb, T. L., Sheeran, P., Miles, E., Niven, K., Wilkinson, I. D., et al. (2014). The neural correlates of regulating another person’s emotions: an exploratory fMRI study. Front. Hum. Neurosci. 8:376. doi: 10.3389/fnhum.2014.00376
PubMed Abstract | CrossRef Full Text | Google Scholar
Hart, H., Radua, J., Nakao, T., Mataix-Cols, D., y Rubia, K. (2013). Meta-análisis de los estudios de imágenes de resonancia magnética funcional de la inhibición y la atención en el trastorno por déficit de atención / hiperactividad: la exploración de la tarea específica, la medicación estimulante, y los efectos de la edad. JAMA Psychiatry. 70, 185-198. doi: 10.1001/jamapsychiatry.2013.277
PubMed Abstract | CrossRef Full Text | Google Scholar
Howard, J. D., y Kahnt, T. (2017). Las representaciones de recompensa específicas de la identidad en la corteza orbitofrontal están moduladas por la devaluación selectiva. J. Neurosci. 37, 3473-3416. doi: 10.1523/JNEUROSCI.3473-16.2017
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, M. E., Budd, T. W., Fulham, W. R., Lancaster, S., Woods, W., Rossell, S. L., et al. (2014). Activación cerebral sostenida que apoya el rendimiento de la tarea de señal de parada. Eur. J. Neurosci. 39, 1363-1369. doi: 10.1111/ejn.12497
PubMed Abstract | CrossRef Full Text | Google Scholar
Hwang, S., White, S. F., Nolan, Z. T., Craig Williams, W., Sinclair, S., y Blair, R. J. (2015). Control de la atención ejecutiva y respuesta emocional en el trastorno por déficit de atención/hiperactividad: un estudio de resonancia magnética funcional. Neuroimage Clin. 9, 545-554. doi: 10.1016/j.nicl.2015.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Ikeda, Y., Okuzumi, H., y Kokubun, M. (2013). Interferencia Stroop/reverse-Stroop en el desarrollo típico y su relación con los síntomas del TDAH. Res. Dev. Disabil. 34, 2391-2398. doi: 10.1016/j.ridd.2013.04.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Karch, S., Thalmeier, T., Lutz, J., Cerovecki, A., Opgen-Rhein, M., Hock, B., et al. (2010). Correlatos neurales (ERP/fMRI) de la selección voluntaria en pacientes adultos con TDAH. Eur. Arch. Psychiatry Clin. Neurosci. 260, 427-440. doi: 10.1007/s00406-009-0089-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaufman, D. A., Keith, C. M., and Perlstein, W. M. (2016). Corteza orbitofrontal y el procesamiento temprano de la novedad visual en el envejecimiento saludable. Front. Aging Neurosci. 8:101. doi: 10.3389/fnagi.2016.00101
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2011). Actividad neuronal que predice la memoria y el olvido posteriores: un meta-análisis de 74 estudios de fMRI. Neuroimage 54, 2446-2461. doi: 10.1016/j.neuroimage.2010.09.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2013). Actividad neuronal diferencial en el reconocimiento de eventos antiguos versus nuevos: un meta-análisis de estimación de probabilidad de activación. Hum. Brain Mapp. 34, 814-836. doi: 10.1002/hbm.21474
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. Y., Wu, T. F., Tsai, J. D., y Yang, E. L. (2016). Aplicabilidad de la tarea de señal de parada para preescolares con TDAH. Percept. Mot. Skills. 123, 162-174. doi: 10.1177/0031512516660715
PubMed Abstract | CrossRef Full Text | Google Scholar
Lenartowicz, A., Verbruggen, F., Logan, G. D., y Poldrack, R. A. (2011). Activación relacionada con la inhibición en la circunvolución frontal inferior derecha en ausencia de señales inhibitorias. J. Cogn. Neurosci. 23, 3388-3399. doi: 10.1162/jocn_a_00031
PubMed Abstract | CrossRef Full Text | Google Scholar
Matthews, S. C., Simmons, A. N., Arce, E., y Paulus, M. P. (2005). Disociación de la inhibición del procesamiento de errores utilizando una tarea inhibitoria paramétrica durante la resonancia magnética funcional. Neuroreport. 16, 755-760. doi: 10.1097/00001756-200505120-00020
PubMed Abstract | CrossRef Full Text | Google Scholar
Mulligan, R. C., Knopik, V. S., Sweet, L. H., Fischer, M., Seidenberg, M., y Rao, S. M. (2011). Correlatos neuronales del control inhibitorio en el trastorno por déficit de atención/hiperactividad en adultos: evidencia de la muestra longitudinal de Milwaukee. Psychiatry Res. 194, 119-129. doi: 10.1016/j.pscychresns.2011.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Nigg, J. T. (1999). El déficit de inhibición de respuesta del TDAH medido por la tarea de parada: replicación con el tipo combinado, la extensión y la calificación del DSM-IV. J. Abnorm. Child Psychol. 27, 393-402. doi: 10.1023/A:1021980002473
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., y Pavuluri, M. N. (2010a). Correlatos neuronales de la inhibición de la respuesta en el trastorno bipolar pediátrico y el trastorno por déficit de atención con hiperactividad. Psychiatry Res. Neuroimaging. 181, 36-43. doi: 10.1016/j.pscychresns.2009.07.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010b). Compromiso diferencial de los sistemas neuronales cognitivos y afectivos en el trastorno bipolar pediátrico y el trastorno por déficit de atención con hiperactividad. J. Int. Neuropsychol. Soc. 16, 106-117. doi: 10.1017/S1355617709991019
PubMed Abstract | CrossRef Full Text | Google Scholar
Pelletier, M. F., Hodgetts, H. M., Lafleur, M. F., Vincent, A., and Tremblay, S. (2016). Vulnerabilidad al efecto de sonido irrelevante en el TDAH adulto. J. Atten. Disord. 20, 306-316. doi: 10.1177/1087054713492563
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollak, Y., Kahana-Vax, G., y Hoofien, D. (2007). Procesos de recuperación en adultos con TDAH: un estudio RAVLT. Dev. Neuropsychol. 33, 62-73. doi: 10.1080/87565640701729789
PubMed Abstract | CrossRef Full Text | Google Scholar
Pretus, C., Picado, M., Ramos-Quiroga, A., Carmona, S., Richarte, V., Fauquet, J., et al. (2016). La presencia de un distractor mejora el rendimiento en la estimación del tiempo en una muestra de adultos con TDAH. J. Atten. Disord. doi: 10.1177/1087054716648776. .
PubMed Abstract | CrossRef Full Text | Google Scholar
Price, J. L. (2007). Definición de la corteza orbital en relación con las conexiones específicas con las estructuras límbicas y viscerales y otras regiones corticales. Annals N.Y. Acad. Sci. 1121, 54-71. doi: 10.1196/annals.1401.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabinak, C. A., MacNamara, A., Kennedy, A. E., Angstadt, M., Stein, M. B., Liberzon, I., et al. (2014). Compromiso prefrontal focal y aberrante durante la regulación de la emoción en veteranos con trastorno de estrés postraumático. Depress. Anxiety. 31, 851-861. doi: 10.1002/da.22243
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, B., Mehler, B., D’Ambrosio, L. A., y Fried, R. (2010). El impacto de las distracciones en los conductores adultos jóvenes con trastorno por déficit de atención e hiperactividad (TDAH). Accid. Anal. Prev. 42, 842-851. doi: 10.1016/j.aap.2009.06.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosch, K. S., Fosco, W. D., Pelham, W. E. Jr., Waxmonsky, J. G., Bubnik, M. G., y Hawk, L. W. Jr. (2016). El refuerzo y la medicación estimulante mejoran la inhibición de respuesta deficiente en niños con trastorno por déficit de atención/hiperactividad. J. Abnorm. Child Psychol. 44, 309-321. doi: 10.1007/s10802-015-0031-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Lim, L., Ecker, C., Halari, R., Giampietro, V., Simmons, A., et al. (2013). Efectos de la edad y el género en las redes neuronales de inhibición de la respuesta motora: de la adolescencia a la mitad de la edad adulta. Neuroimage. 83, 690-703. doi: 10.1016/j.neuroimage.2013.06.078
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Oosterlaan, J., Sergeant, J. A., Brandeis, D., y Leeuwen, T. V. (1998). Disfunción inhibitoria en niños hiperactivos. Behavior. Brain Res. 94, 25-32. doi: 10.1016/S0166-4328(97)00166-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Shalom, D. B. (2009). La corteza prefrontal medial y la integración en el autismo. Neuroscientist. 15, 589-598. doi: 10.1177/1073858409336371
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, J. L., Jamadar, S., Provost, A. L., y Michie, P. T. (2013). Inhibición motora y no motora en la tarea Go/NoGo: un estudio de ERP y fMRI. Int. J. Psychophysiol. 87, 244-253. doi: 10.1016/j.ijpsycho.2012.07.185
PubMed Abstract | CrossRef Full Text | Google Scholar
Soliman, A. M., y Elfar, R. M. (2017). Falsa memoria en adultos con TDAH: una comparación entre subtipos y controles normales. J. Attent. Disord. 21, 986-996. doi: 10.1177/1087054714556814
PubMed Abstract | CrossRef Full Text | Google Scholar
Sørensen, L., Plessen, K. J., Adolfsdottir, S., y Lundervold, A. J. (2014). La especificidad de la puntuación de interferencia de errores Stroop para el TDAH en los niños. Child Neuropsychol. 20, 677-691. doi: 10.1080/09297049.2013.855716
PubMed Abstract | CrossRef Full Text | Google Scholar
Soto, D., Humphreys, G. W., y Rotshtein, P. (2007). Disociando los mecanismos neurales de la guía de selección visual basada en la memoria. Proc. Natl. Acad. Sci. U.S.A. 104, 17186-17191. doi: 10.1073/pnas.0703706104
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinelli, S., Joel, S., Nelson, T. E., Vasa, R. A., Pekar, J. J., y Mostofsky, S. H. (2011). Diferentes patrones neuronales se asocian con los ensayos que preceden a los errores de inhibición en los niños con y sin trastorno por déficit de atención/hiperactividad. J. Am. Acad. Child Adolesc. Psychiatry. 50, 705-715. doi: 10.1016/j.jaac.2011.03.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsujimoto, S., Yasumura, A., Yamashita, Y., Torii, M., Kaga, M., and Inagaki, M. (2013). Aumento de la oxigenación prefrontal relacionada con la memoria de trabajo resistente a los distractores en niños con trastorno por déficit de atención/hiperactividad (TDAH). Child Psychiatry Hum. Dev. 44, 678-688. doi: 10.1007/s10578-013-0361-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcott, C. M., y Landau, S. (2004). La relación entre la desinhibición y la regulación de la emoción en los niños con trastorno por déficit de atención e hiperactividad. J. Clin. Child Adolesc. Psychol. 33, 772-782. doi: 10.1207/s15374424jccp3304_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Xiong, K., Zhang, Y., Qiu, M., Zhang, J., Sang, L., Wang, L., et al. (2013). Regulación de la emoción negativa en pacientes con trastorno de estrés postraumático. PLoS ONE 8:e81957. doi: 10.1371/journal.pone.0081957
PubMed Abstract | CrossRef Full Text | Google Scholar
Yarmolovsky, J., Szwarc, T., Schwartz, M., Tirosh, E., y Geva, R. (2016). Control ejecutivo caliente y respuesta a un estimulante en un ensayo aleatorio doble ciego en niños con TDAH. Eur. Arch. Psychiatry Clin. Neurosci. 267, 73-82. doi: 10.1007/s00406-016-0683-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Yasumura, A., Kokubo, N., Yamamoto, H., Yasumura, Y., Nakagawa, E., Kaga, M., et al. (2014). Evaluación neuroconductual y hemodinámica de la interferencia Stroop y Stroop inversa en niños con trastorno por déficit de atención/hiperactividad. Brain Dev. 36, 97-106. doi: 10.1016/j.braindev.2013.01.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Zsuga, J., Biro, K., Tajti, G., Szilasi, M. E., Papp, C., Juhasz, B., et al. (2016). Uso ‘proactivo’ de la congruencia pista-contexto para construir la función de recompensa del aprendizaje por refuerzo. BMC Neurosci. 17:70. doi: 10.1186/s12868-016-0302-7
PubMed Abstract | CrossRef Full Text | Google Scholar