Eukaryooteilla on kaksi pääasiallista solunjakautumistyyppiä: mitoosi ja meioosi. Mitoosia käytetään uusien kehon solujen tuottamiseen kasvua ja paranemista varten, kun taas meioosia käytetään sukupuolisolujen (munasolujen ja siittiöiden) tuottamiseen. Meioosia käsitellään myöhemmässä luvussa.
Solusykli on järjestetty tapahtumasarja, johon liittyy solun kasvu ja solunjakautuminen, joka tuottaa mitoosin avulla kaksi uutta tytärsolua. Solusyklin pituus vaihtelee suuresti jopa yksittäisen organismin solujen sisällä. Ihmisillä solujen vaihtumistiheys vaihtelee alkion varhaiskehityksen muutamasta tunnista epiteelisolujen keskimäärin kahdesta viiteen vuorokauteen tai koko ihmisen elinikään, joka kuluu jakautumatta erikoistuneissa soluissa, kuten aivokuoren neuroneissa tai sydänlihassoluissa. Myös aika, jonka solu viettää kussakin solusyklin vaiheessa, vaihtelee. Kun nopeasti jakautuvia nisäkässoluja kasvatetaan viljelyssä (elimistön ulkopuolella optimaalisissa kasvuolosuhteissa), syklin pituus on noin 24 tuntia. Solusyklin tapahtumien ajoitusta säätelevät sekä solun sisäiset että solun ulkopuoliset mekanismit.
Solut kulkevat solunjakautumisen tiellä läpi sarjan tarkasti ajoitettuja ja tarkoin säädeltyjä kasvun, DNA:n replikaation ja jakautumisen vaiheita, jotka tuottavat kaksi geneettisesti identtistä solua. Solusyklissä on kaksi päävaihetta: interfaasi ja mitoosivaihe (kuva 1). Interfaasin aikana solu kasvaa ja DNA:ta monistetaan. Mitoosivaiheen aikana replikoitunut DNA ja sytoplasman sisältö erkanevat toisistaan ja solu jakautuu.
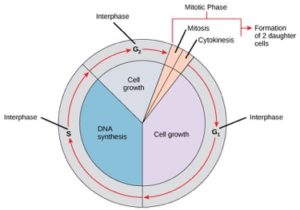
Välivaiheen aikana solu käy läpi normaaleja prosesseja ja valmistautuu samalla solunjakautumiseen. Jotta solu voi siirtyä interfaasista mitoosivaiheeseen, monien sisäisten ja ulkoisten ehtojen on täytyttävä. Interfaasin kolmea vaihetta kutsutaan G1-, S- ja G2-vaiheiksi.
G1-vaihe (ensimmäinen aukko)
Interfaasin ensimmäistä vaihetta kutsutaan G1-vaiheeksi (ensimmäinen aukko), koska mikroskooppiselta kannalta katsottuna vain vähän muutoksia on näkyvissä. G1-vaiheen aikana solu on kuitenkin biokemiallisella tasolla varsin aktiivinen. Solu kerryttää kromosomaalisen DNA:n ja siihen liittyvien proteiinien rakennuspalikoita sekä kerryttää riittävästi energiavarastoja, jotta se voi suorittaa kunkin kromosomin monistamistehtävän tumaan.
S-vaihe (DNA:n synteesi)
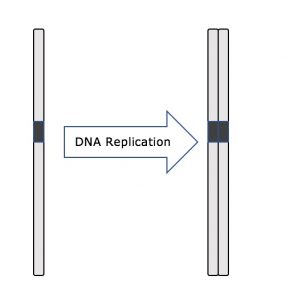
Välivaiheen aikana ydin-DNA säilyy puolikondensoituneessa kromatiinimuodossa. S-vaiheessa DNA:n replikaatio voi edetä mekanismeilla, jotka johtavat identtisten DNA-molekyyliparien – sisarkromatidien – muodostumiseen, jotka kiinnittyvät kiinteästi sentromeeriselle alueelle (kuva 2).
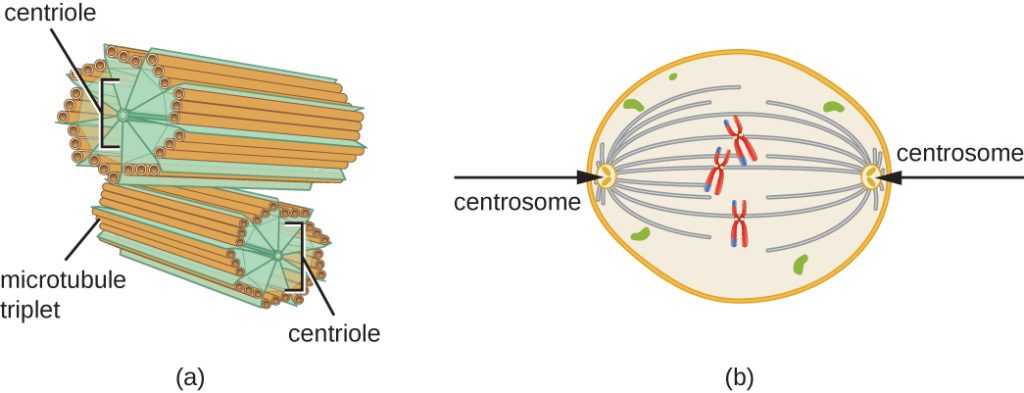
S-vaiheen aikana myös sentrosomi monistuu. Kahdesta sentrosomista syntyy mitoottinen kara, laite, joka organisoi kromosomien liikkumista mitoosin aikana. Eläinsolujen sentrosomeihin liittyy kunkin eläinsolun keskellä pari sauvamaista kohdetta, sentrioleja, jotka ovat suorassa kulmassa toisiinsa nähden. Sentriolit auttavat organisoimaan solunjakautumista. Sentrioleja ei ole muiden eukaryoottilajien, kuten kasvien ja useimpien sienten, sentrosomeissa.
G2-vaihe (toinen aukko)
G2-vaiheessa solu täydentää energiavarastojaan ja syntetisoi kromosomien manipulointiin tarvittavia proteiineja. Joitakin solun organelleja monistetaan, ja sytoskeletti puretaan, jotta saadaan resursseja mitoosivaihetta varten. G2-vaiheen aikana voi tapahtua solun lisäkasvua. Viimeiset valmistelut mitoosivaihetta varten on saatava päätökseen, ennen kuin solu voi siirtyä mitoosin ensimmäiseen vaiheeseen.
Mitoosivaihe
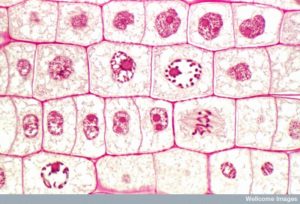
Kahden tytärsolun muodostamiseksi tuman ja sytoplasman sisältö on jaettava. Mitoosivaihe on monivaiheinen prosessi, jonka aikana monistuneet kromosomit kohdistetaan, erotetaan ja siirretään solun vastakkaisiin napoihin, minkä jälkeen solu jakautuu kahdeksi uudeksi identtiseksi tytärsoluksi. Mitoosivaiheen ensimmäinen osa, mitoosi, koostuu viidestä vaiheesta, jotka toteuttavat ydinjakautumisen (kuva 5). Mitoosivaiheen toinen osa, jota kutsutaan sytokinesikseksi, on sytoplasman osien fyysinen erottaminen kahdeksi tytärsoluksi. Vaikka mitoosin vaiheet ovat samankaltaisia useimmilla eukaryooteilla, sytokinesisprosessi on aivan erilainen eukaryooteilla, joilla on soluseinät, kuten kasvisoluilla.
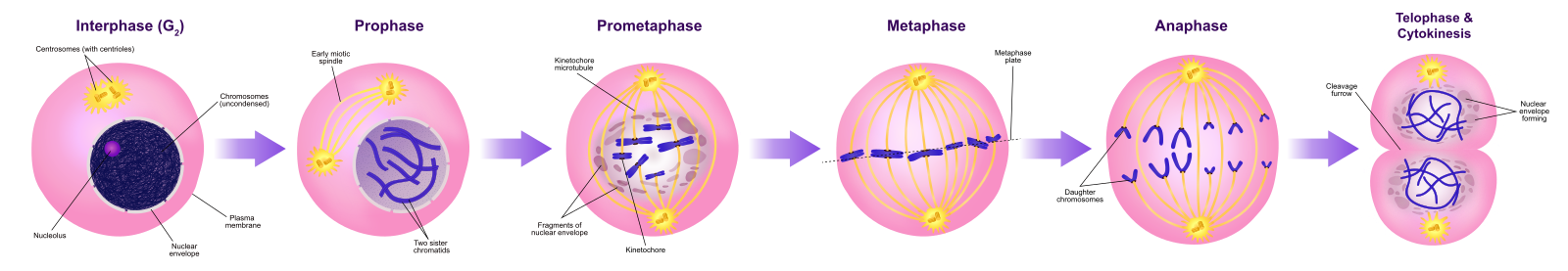
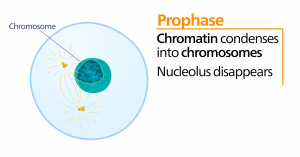
Profaasi
Profaasin eli ”ensimmäisen vaiheen” aikana ydinkuori alkaa dissosioitua pieniksi rakkuloiksi, ja kalvomaiset organellit (kuten Golgin apparaatti ja endoplasminen retikulum), pirstoutuvat ja hajaantuvat kohti solun reunoja. Nukleoli häviää. Sentrosomit alkavat siirtyä solun vastakkaisiin napoihin. Mikrotubulukset, jotka muodostavat mitoottisen karan, ulottuvat sentrosomien väliin ja työntävät niitä kauemmas toisistaan mikrotubulussäikeiden pidentyessä. Sisaruskromatidit alkavat kietoutua tiiviimmin kondensiiniproteiinien avulla, ja ne tulevat näkyviin valomikroskoopissa.
Prometafaasi
Prometafaasin eli ”ensimmäisen muutosvaiheen” aikana monet profaasissa aloitetut prosessit jatkavat etenemistään. Ydinkuoren jäänteet pirstoutuvat. Mitoottinen kara jatkaa kehittymistään, kun lisää mikrotubuluksia kerääntyy ja venyy entisen ydinalueen pituudelle. Kromosomit tiivistyvät ja erilaistuvat. Kullekin sisarkromatidille kehittyy proteiinirakenne nimeltä kinetokori sentromerialueella.
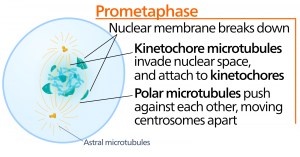
Kinetokorin proteiinit vetävät puoleensa ja sitovat mitoottisen karan mikrotubuluksia. Kun karan mikrotubulukset ulottuvat sentrosomeista, osa näistä mikrotubuluksista joutuu kosketuksiin kinetokorien kanssa ja sitoutuu niihin lujasti. Kun mitoottinen säie kiinnittyy kromosomiin, kromosomi suuntautuu, kunnes sisarkromatidien kinetokuoret ovat vastakkaisia napoja vasten. Lopulta kaikki sisarkromatidit kiinnittyvät kinetokoriensa kautta vastakkaisten napojen mikrotubuluksiin. Karan mikrotubuluksia, jotka eivät kiinnity kromosomeihin, kutsutaan polaarisiksi mikrotubuluksiksi. Nämä mikrotubulukset limittyvät toisiinsa kahden navan puolivälissä ja edistävät solun pidentymistä. Astraaliset mikrotubulukset sijaitsevat lähellä napoja, auttavat karan suuntautumisessa ja niitä tarvitaan mitoosin säätelyyn.

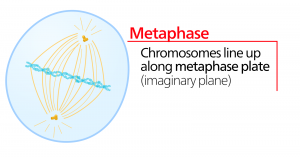
Metafaasi
Metafaasissa, ”muutosvaiheessa”, kaikki kromosomit ovat kohdakkain tasossa, jota kutsutaan nimellä metafaasilevy eli ekvatoriaalitaso, joka on solun molempien napojen keskellä. Sisaruskromatidit ovat edelleen tiukasti kiinni toisissaan kohesiiniproteiineilla. Tällä hetkellä kromosomit ovat maksimaalisesti tiivistyneitä.
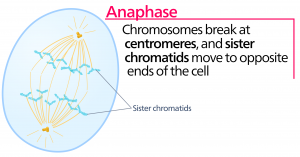
Anaphaasi
Anafaasin eli ”ylöspäin suuntautuvan vaiheen” aikana kohesiiniproteiinit hajoavat ja sisarkromatidit erkanevat sentromeerin kohdalla. Kukin kromatidi, jota nyt kutsutaan kromosomiksi, vedetään nopeasti kohti sentrosomia, johon sen mikrotubulus on kiinnittynyt. Solusta tulee näkyvästi pitkänomainen (soikea), kun polaariset mikrotubulukset liukuvat toisiaan vasten metafaasilevyllä, jossa ne ovat päällekkäin.
Telopaasi
Telopaasin eli ”etäisyysvaiheen” aikana kromosomit saavuttavat vastakkaiset navat ja alkavat dekondensoitua (purkautua) rentoutuen kromatiinikokoonpanoon. Mitoottiset spindelit depolymerisoituvat tubuliinimonomeereiksi, joita käytetään kunkin tytärsolun sytoskelettikomponenttien kokoamiseen. Ydinkuori muodostuu kromosomien ympärille, ja nukleosomit ilmestyvät ydinalueelle.
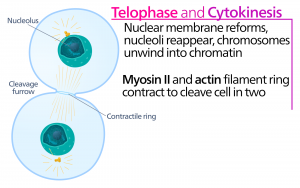
Sytokinesis
Sytokinesis eli ”soluliike” on mitoosivaiheen toinen päävaihe, jonka aikana solunjakautuminen saatetaan päätökseen sytoplasman osien fyysisen erottumisen kautta kahdeksi tytärsoluksi. Jakautuminen on päättynyt vasta, kun solun osat ovat jakautuneet ja erkaantuneet kokonaan kahdeksi tytärsoluksi. Vaikka mitoosin vaiheet ovat samankaltaisia useimmilla eukaryooteilla, sytokinesisprosessi on aivan erilainen eukaryooteilla, joilla on soluseinät, kuten kasvisoluilla.
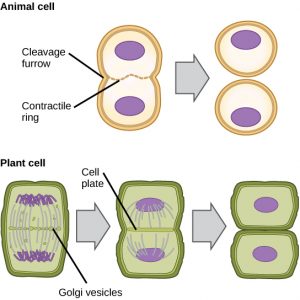
Eläinsolujen kaltaisissa soluissa, joilla ei ole soluseiniä, sytokinesis seuraa anafaasin alkamista. Aktiinifilamenteista koostuva supistuva rengas muodostuu juuri plasmakalvon sisäpuolelle entisen metafaasilevyn kohdalle (kuva 12). Aktiinifilamentit vetävät solun päiväntasaajaa sisäänpäin muodostaen halkeaman. Tätä halkeamaa tai ”säröä” kutsutaan halkaisukouruksi. Aktiinirenkaan supistuessa uurre syvenee, ja lopulta kalvo halkeaa kahtia.
Kasvisoluissa tytärsolujen väliin on muodostettava uusi soluseinämä. Interfaasin aikana Golgin laitteeseen kertyy entsyymejä, rakenneproteiineja ja glukoosimolekyylejä ennen kuin se hajoaa vesikkeleiksi ja leviää koko jakautuvaan soluun (kuva 12). Telofaasin aikana nämä Golgin vesikkelit kulkeutuvat mikrotubulusten varassa muodostaen metafaasilevyn kohdalla fragmoplastin (vesikkelirakenteen). Siellä vesikkelit sulautuvat ja yhdistyvät keskeltä kohti soluseinämiä; tätä rakennetta kutsutaan solulevyksi. Kun lisää vesikkeleitä sulautuu, solulevy laajenee, kunnes se sulautuu solun seinämiin solun reuna-alueella. Entsyymit käyttävät kalvokerrosten väliin kertynyttä glukoosia uuden soluseinän rakentamiseen. Golgin kalvoista tulee plasmakalvon osia uuden soluseinän molemmin puolin.
Yhteenveto mitoosista ja sytokinesiksestä
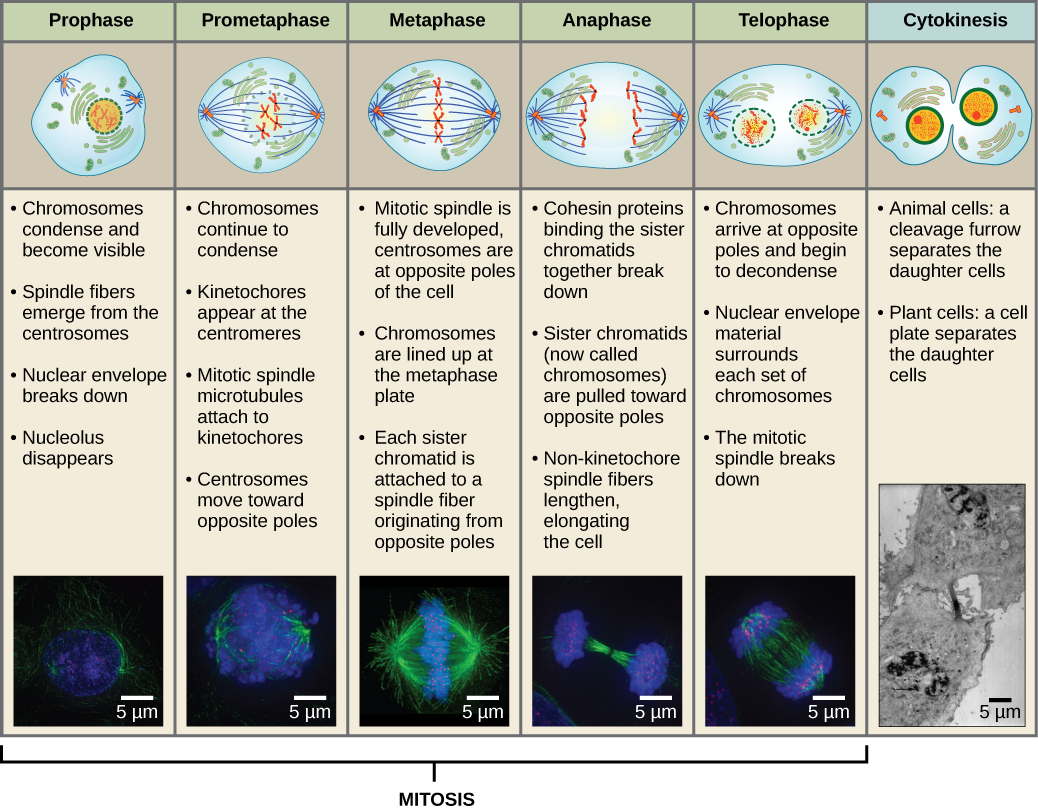
Kuvio 13 Mitoosi jaetaan viiteen vaiheeseen – profaasiin, prometafaasiin, metafaasiin, anafaasiin ja telopaasiin. Alareunan kuvat on otettu fluoresenssimikroskopialla soluista, jotka on keinotekoisesti värjätty fluoresoivilla väriaineilla: sininen fluoresenssi osoittaa DNA:ta (kromosomit) ja vihreä fluoresenssi osoittaa mikrotubuluksia (karalaite). (luotto ”mitoosipiirrokset”: Mariana Ruiz Villarealin työn muokkaus; luotto ”mikrokuvat”: Roy van Heesbeenin työn muokkaus; luotto ”sytokinesismikrokuva”: Wadsworth Center/New York State Department of Health; scale-bar data from Matt Russell)
G0 Phase
Eivät kaikki solut noudata klassista solusyklimallia, jossa vastaperustettu tyttärisolu siirtyy välittömästi interfaasiin, jota seuraa välittömästi mitoosivaihe. G0-vaiheessa olevat solut eivät aktiivisesti valmistaudu jakautumaan. Solu on rauhallisessa (inaktiivisessa) vaiheessa, kun se on poistunut solusyklistä. Jotkin solut siirtyvät G0-vaiheeseen väliaikaisesti, kunnes ulkoinen signaali käynnistää G1-vaiheen alkamisen. Muut solut, jotka eivät koskaan tai harvoin jakaudu, kuten kypsät sydänlihas- ja hermosolut, pysyvät G0-vaiheessa pysyvästi).