
















































Tämä kuvitus on saatavana tulostettavana mustavalkoisena värityskuvana. Klikkaa tästä ja napsauta hiiren oikealla painikkeella kuvaa, joka avautuu uuteen ikkunaan ja tallenna tietokoneellesi.
Nimi: C.megalodon(Isohammas).
Fonetiikka: Meg-ah-low-don.
Nimimerkki: Meg-ah-low-don.
Nimimerkki: Meg-ah-low-don: Louis Agassiz – 1843.
Synonyymit: Procarcharodon megalodon,Megaselachus megalodon.
Luokitus: Chordata, Chondrichthyes,Elasmobranchii, Lamniformes.
Lajit: Carcharodon megalodon,Carcharocles megalodon, Procarcharodonmegalodon tai jopa Otodus megalodon katso tekstistä täydellinen selitys.
Ruokavalio:
Koko:
Koko: Monia arvioita, mutta yleensä 15-17 metriä pitkä.
Tunnetut sijainnit:
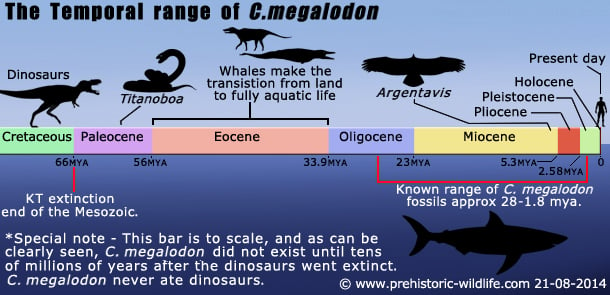
Ajanjakso: Oligoseenin loppupuolelta pleistoseenin alkupuolelle.
Fossiilinen edustus: Enimmäkseen hampaita, mutta myös joitakin nikamia tunnetaan.
Hampaat ja nikamat – Thefossils ofmegalodon
Hampaat ovat ylivoimaisesti yleisimpiä C. megalodonin jäännöksiä, ja suuremmat hampaat lähestyvät 18 senttimetrin pituutta. Ei ole varmaa, kuinka kauan ihmiset ovat keränneet C. megalodonin hampaita, mutta vasta vuonna 1667 tiede tunnusti ne haihampaiksi. Ennen tätä ajankohtaa ihmiset luulivat yleensäkin ja vilpittömin mielin, että C. megalodonin hampaita oli C. megalodon.Totuus niistä selvisi, kun tanskalainen luonnontieteilijä Nicolas Steno tunnisti oikein yhden näistä ”lohikäärmeen kielistä” ja mainitsi sen kirjassaan The Head of a SharkDissected.
C.megalodonin hampaita verrataan tavallisesti valkohain hampaisiin, koska ne ovat pinnallisesti samankaltaisia: ne ovat kolmionmuotoisia ja hammastettuja.Tämän samankaltaisuuden, erityisesti hammastuksen, vuoksi ajatellaan, että C.megalodonilla oli sama puremistapa kuin valkohailla. Tämä tarkoittaisi sitä, että C.megalodon puree saaliinsa ja ravistelee sitten päätään puolelta toiselle niin, että hampaiden hammastukset sahaavat lihan läpi.
On jopa löydetty lähes täydellisiä C.megalodonin hampaiden sarjoja samalta yksilöltä, jotka on löydetty yhdessä.Nämä paljastavat, että C.megalodon oli muiden haiden kaltainen siinä mielessä, että suurimmat hampaat olivat suun etupuolella. Mitä kauempana etupuolella hampaat olivat, sitä tasaisemman kolmionmuotoiset ne olivat. Mitä enemmän hampaat menivät sivulle leuan takaosaan, sitä sisäänpäin kaarevammiksi ne tulivat.Nämä hampaat koukistuivat saaliin lihaan, mikä vaikeutti saaliin pakenemista etuhampaiden leikatessa sitä.
Yksi yleisimmistä myytteistä, joita C. megalodonista useimmiten kerrotaan, on se, että se tunnetaan vain hampaiden perusteella. Todellisuudessa hampaat ovat ylivoimaisesti yleisimpiä fossiileja C. megalodonista, mutta myös selkärankaa on saatu talteen pieni ja kasvava määrä. Hain biologiassa yleensä,luuranko koostuu aina ”pehmeästä” rustosta, mutta selkäranka koostuu kovemmasta kalkkiutuneesta rustosta.Tämä tarkoittaa sitä, että vaikka nikamat voivat hajota ja usein hajoavatkin, ne voivat säilyä pidempään, mikä lisää niiden fossiilisoitumisen mahdollisuutta.

Koko – Megalodonin ylemmän koon arvioiminen
Mikäli hampaat ovat C. megalodonin todettiin olevan haiden hampaita,useimmat ihmiset kysyivät heti, ”mutta kuinka suuri hai oli?”. Rehellinen vastaus on, ettemme tarkalleen tiedä varmasti, koskaei ole olemassa täydellistä yksilöä, johon joku voisi ottaa mittanauhan.Sen sijaan paleontologit ja iktyologit joutuvat käyttämään sitä, mitä heillä on, tässä tapauksessa hampaita. Hain pituuden arvioimisesta hampaiden analyysin perusteella on tehty monia tutkimuksia, joista neljä yleisimmin mainittua on tässä.
C. megalodonin pituuden arvioimiseksi käytettiin aikoinaan menetelmää, jossa mitattiin hammaskiilteen korkeus. Menetelmän kehittivät John E.Randall vuonna 1973, ja sen tuloksena saatiin arvioksi 13 metriä.Laajempi tiedeyhteisö ei kuitenkaan pidä menetelmää täsmällisenä, koska menetelmä syntyi ensin vertaamalla valkohain hampaita, ja vaikka nämä hampaat näyttävätkin päällisin puolin samankaltaisilta, ne eroavat itse asiassa huomattavasti C. megalodonin hampaista.Lisäksi hammaskiilteen määrä voi vaihdella jopa saman lajin hampaiden välillä kulumisen ja huonon säilyvyyden vuoksi.
Mikäli Michael D. Gottfriedin, Leonard J. V. Compagnon ja S. Curtis Bowmanin vuonna 1996 kehittämä arviointimenetelmä perustuu hampaan vinon korkeuden ja hain pituuden välisen korrelaation määrittämiseen.Viistokorkeus ei ole kokonaiskorkeus vaan pituus hampaan kärjestä sivureunaan, ja se on hampaan todellinen leikkauspituus. Tämäkin tutkimus perustui valkohampaiden tutkimukseen, ja sen tuloksena saatiin arvioksi 15,9 metriä. Vaikka tämä menetelmä olisikin luotettava, se voi kuitenkin antaa arvioita vain tietyistä hampaista eikä välttämättä lajista.Clifford Jeremiah kehitti toisen menetelmän, joka perustuu hampaan juuren leveyteen, joka on kruunun alapuolella oleva osa, jota liha peittää. Kyseessä on yksinkertainen periaate, jonka mukaan hampaan kruunun leveyden perusteella voidaan laskea leuan leveys, jota puolestaan voidaan käyttää hain pituuden laskemiseen. Yksinkertaisimmillaan menetelmä perustuu siihen, että hampaan juurta on yksi senttimetri jokaista 129 senttimetriä kohti.Näin saatiin 15,5 metrin pituusarvio hampaalle, jossa oli noin 12 senttimetriä juurta.
Tohtori Kenshu Shimada kehitti vuonna 2002 toisen, monimutkaisemman menetelmän, joka perustuu hampaan kruunun mittasuhteisiin. Kun tätä menetelmää käytettiin hampaaseen, jota Gottfried ym. olivat käyttäneet vuonna 1996, pituusarvioksi saatiin 15,1 metriä.Suurin koskaan tällä menetelmällä saatu arvio saatiin Panamasta peräisin olevasta erityisen suuresta hampaasta, jonka ruumiinpituudeksi saatiin 16,8 metriä.
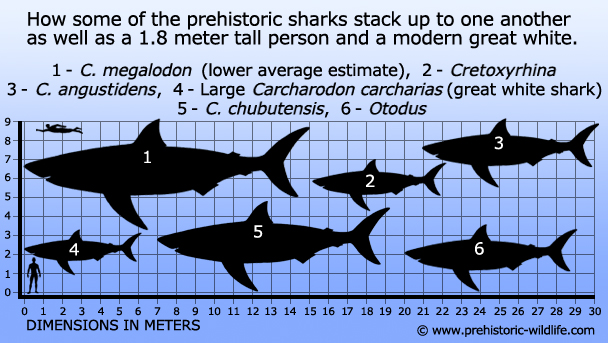
Arvioidun ruumiinpituuden vaihteluväliä 15-16 metriä pidetään melko turvallisena C. megalodonin osalta, ja jopa 17 metrin pituisia estimaatteja pidetään realistisesti mahdollisina.Suurempia estimaatteja on olemassa, ja on esitetty väitteitä, joiden mukaan C. megalodon olisi voinut saavuttaa 17 metriä. megalodon voisi lähestyä ja jopa ylittää 20 metrin pituuden, mutta suurinta osaa näistä arvioista pidetään vain mahdollisuuksina, eivätkä ne vastaa nykyisiä fossiilisia todisteita. Silti vaatimattomampikin 15-metrinen C. megalodon olisi kääpiöitynyt minkä tahansa muun elävän tai sukupuuttoon kuolleen hain rinnalla, ja jopa valkohai kalpenisi sen jättiläismäisen koon rinnalla.
Miten paljon C. megalodon olisi voinut kasvaa? C. megalodon painoi, on toinen tutkimuskysymys, vaikka sen yksi koko voi tuottaa vieläkin laajempia arvioita. Nykyiset arviot perustuvat jälleen kerran vertailuun valkohain kanssa, ja 15,9-metrisen C. megalodonin arvioksi on arvioitu 47 tonnia. 17-metrisen C. megalodonin arvioksi on arvioitu 59 tonnia ja 20,3-metrisen C. megalodonin arvioksi 103 tonnia.Kuten olette ehkä huomanneet, C. megalodonin paino ei kasva tasaisesti pituuden myötä, minkä vuoksi 20,3-metrinen hai on noin kolmanneksen pidempi kuin 15,9-metrinen hai, mutta painaa silti yli kaksinkertaisesti hieman pienempään yksilöön verrattuna. Tämä johtuu siitä, että hai ei ole pelkästään pidempi, vaan se on myös suhteessa paksumpi ja järeämpi, ja sen lihakset ovat kasvaneet suuremman rungon liikuttamiseksi. Tämä tarkoittaa myös sitä, että eläin, olipa se sitten hai tai mikä tahansa muu eläin, saavuttaa aina koon, josta se ei voi enää kasvaa, koska sen elinympäristö ei kestä ravintoa, mikä on luonnollinen tosiasia, joka perustuu siihen logiikkaan, että isompi ruumis tarvitsee enemmän ravintoa saadakseen energiaa polttoaineekseen.
Mahdollisesti biologia – Megalodonin sisäinen toiminta
Suuren ruumiinkoonsa vuoksi C. megalodonin uskotaan lähes varmasti eläneen jättilämmön vaikutuksesta. Tällöin olento on niin massiivinen, että sen keho pitää lämpöä sisällään niin, että lihaksen ja lihan uloimmat kerrokset itse asiassa eristävät sisäelimet ympäristöstä. Tämä aiheuttaa aineenvaihdunnan kiihtymisen, mikä puolestaan tekee olennosta aktiivisemman. C. megalodon on saattanut viedä jättilämpöprosessia vielä pidemmälle ohjaamalla lämpimämmän veren virtauksen päähänsä ja kuonoonsa, kuten valkohain tiedetään tekevän. Tämä saisi C. megalodonin aivot ja aistielimet, kuten haju- ja näköaistit sekä sähköhermoa havaitsevat ampullat, toimimaan lämminverisellä aineenvaihduntatasolla.C. megalodonin hampaita tunnetaan jokaiselta mantereelta, mikä todistaa, että sen levinneisyysalue on ollut kosmopoliittinen, kuten monilla muillakin menestyksekkäillä merten saalistajilla.C. megalodonin levinneisyys näyttää kuitenkin vähentyneen valtamerten jäähtyessä, erityisesti plioseenin loppupuolella.
Yleisilmeeltään C. megalodon on todennäköisesti ollut hyvin samankaltainen kuin nykyään meressä uivat lamellihait,
vaikkakin suuren kokonsa ja luontaisen voimansa vuoksi se on todennäköisesti ollut melko tukevarakenteinen nykyaikaisiin haihin verrattuna.Myös ruston luuranko olisi todennäköisesti ollut kestävämpi, jotta se olisi kestänyt suurempien lihasten ja voimakkaampien saaliseläinten aiheuttamat suuremmat rasitukset.
C.megalodonilla oli todennäköisesti liikkuva yläleuka, joka liikkui ylös- ja alaspäin ja jota voitiin työntää eteenpäin erillään muusta kehosta.Tällaista leukaa näkee nykyaikaisissa haikaloissa, ja sen vuoksi hai, joka on aikeissa pureutua johonkin, näyttää hyvin erilaiselta kuin hai, joka vain risteilee.Kun yläleuka työntyy eteenpäin, ylähampaat ja leuka tulevat näkyviin, kun taas tavallisesti suu peittää ne. Tällä tavoin liikkuvan leuan ansiosta C. megalodon tarttui saaliiseensa hitaamman leukansa pitäessä sitä paikoillaan ja ravisteli sitten päätään puolelta toiselle niin, että hampaiden viillot leikkasivat palan lihaa irti.Tämän jälkeen yläleuka vedetään takaisin, jolloin lihakappale vedetään suuhun. Hai toistaa tämän prosessin, kunnes se on syönyt tarpeekseen.
Hampaiden löytymiseen välittömästi liittyvä tutkimusalue on C. megalodonin puremisvoiman arviointi. Tämä on tärkeää tietää, sillä se, miten voimakkaasti eläin pystyy puremaan jotain, voi antaa viitteitä siitä, millaista saalista eläin söi ja miten se söi sen.Vuonna 2008 luotiin biomekaaninen tietokonemalli valkohaiden purentavoiman arvioimiseksi. Kun tätä mallia sovellettiin C. megalodoniin, paljastui, että 15,9-metrinen C. megalodon, joka on keskimäärin suuri, kykeni käyttämään 108 514 newtonin purentavoimaa, mikä on hieman yli 11 tonnia.Kun arvioitiin 20,3 metrin pituisen C. megalodonin, joka on C. megalodonin potentiaalisen kokoskaalan yläpää, purentavoima, tulokseksi saatiin 182 201 newtonia eli yli 18,5 tonnia. Tämä tarkoittaa, että jopa alhaisemmat arviot C. megalodonin purentavoimasta olivat yli 18,5 tonnia. megalodonin puremavoimat ovat silti paljon suuremmat kuin julmalla istukkahain Dunkleosteuksella ja jopa suuremmat kuin mahtavalla Tyrannosaurusrexillä. Toinen huomioon otettava seikka on se, että C. megalodon todennäköisesti ravisteli päätään puolelta toiselle kuten muutkin hait, joilla oli samanlainen hampaisto. Tämä tarkoitti sitä, että C. megalodonin epäonniseen saaliiseen kohdistuneet todelliset voimat olivat todennäköisesti vieläkin suuremmat.
Mikäli useimmat C. megalodonia koskevat tutkimukset eivät ole osoittaneet, että C. megalodonia olisi voitu tappaa. megalodonin hampaisiin, eräs alue, joka jää usein huomiotta, ovat nikamat. Tunnettujen nikamien pienen määrän vuoksi kaikki eivät pysty tutkimaan niitä hampaiden tapaan, mutta yksi asia, joka nikamissa on selvästi nähtävissä, ovat konsentriset renkaat. Nämä konsentriset renkaat ovat pohjimmiltaan samoja kuin puun kannossa näkyvät kasvurenkaat, ja ne näkyvät myös muissa nykyaikaisissa haikaloissa, jotka uivat nykyään valtamerissä.Näiden samankeskisten renkaiden laskemisen perusteella fossiileista tunnettujen C. megalodonien iäksi on arvioitu 25-40 vuotta, ja paleontologit arvelevat, että C. megalodonit ovat saattaneet elää tätä pidempäänkin.
Nuoret megalodonit – poikaset ja kasvualustat
Nuoret megalodonit – poikaset ja kasvualustat
Suuren kokonsa ja pelagisen elämäntyylinsä vuoksi C. megalodonit ovat voineet elää paljon kauemmin. C. megalodonin uskotaan synnyttäneen eläviä poikasia. Siitä, miten C. megalodonit tarkalleen ottaen synnyttivät, voidaan kuitenkin vielä kiistellä, sillä hailla on kaksi tapaa tehdä tämä. Ensimmäistä kutsutaan vivipariteetiksi, ja siinä poikanen, eli hainpoikanen, kasvaa emon sisällä, kunnes se on valmis syntymään. Näin syntyneet hainpoikaset ruokitaan ravintoaineilla napanuoran kautta, ja kun poikanen on synnytetty, istukka yleensä poistuu heti sen jälkeen.
Kakkosmenetelmä on ovoviviparia, jossa poikanen kehittyy munan sisällä. Ovovivipariaa harjoittavilla hailla munaa ei kuitenkaan lasketa, vaan se säilyy emohain sisällä. Näiden hainpoikasten poikaset eivät ole napanuoralla kiinni emossaan, vaan ne käyttävät munan keltuaista kehityksensä aikana.Molemmissa kehitystyypeissä poikaset ovat tajuissaan ja kykenevät uimaan omin voimin, kun ne poistuvat emonsa kehosta. C.megalodonin poikaset kuvataan yleensä pyrstö edellä, ja vaikka tätä pidetäänkin tavanomaisena tapana, jotkin nykyiset hailajit syntyvät itse asiassa pää edellä.
Samoin kuin nykyisissä haikaloissa, C. megalodonin poikaset syntyvät myös nykyisin. megalodon ei todennäköisesti synnyttänyt poikasiaan missä tahansa, vaan valitsi niin sanotut kasvattamoalueet. Kasvattamoalue on paikka, jossa nuori hai voi elää ja metsästää turvassa muilta suuremmilta saalistajilta. Kasvattamoalueilla eläviä haita pidetään generalisteina, jotka hyökkäävät ja syövät kaikenlaista, mukaan lukien kaloja, pääjalkaisia (kuten mustekaloja ja seepioita), kilpikonnia ja melkein mitä tahansa muuta, minkä ne voivat napata.
Tunnistaakseen mahdolliset kasvualueet paleontologit etsivät pienempien C. megalodonin hampaiden keskittymiä. Yksi alue, jolla näitä hampaita näyttäisi olevan runsaasti, on Keski-Amerikka ja Yhdysvaltojen eteläisimmät alueet. Oligoseenikaudella Panamaa ei ollut olemassa, koska silloiset korkeammat merenpinnat upottivat suuren osan alueesta veden alle. Tämä alue tunnettiin nimellä Keski-Amerikan meriväylä, ja se muodosti Tyynenmeren ja Atlantin välisen valtameren solan.Alueella oli todennäköisesti valtavasti matalia vesialueita, jotka eivät yksinkertaisesti olleet tarpeeksi syviä isommille petoeläimille, joten ne olivat suhteellisen turvallisia paljon pienemmille C. megalodonin poikasille.

Fossiiliset todisteet viittaavat vahvasti siihen, että Pohjois- ja Etelä-Amerikan välisillä matalilla vesialueilla oli mioseeniaikana hyvin suuri määrä nuoria C. megalodonin poikasia. Fossiiliesiintymät viittaavat myös siihen, että varhaiset valaslajit, kuten valaat (violetti), käyttivät tuolloin Keski-Amerikan meriväylänä tunnettua aluetta Tyynenmeren ja Atlantin vesien välisenä kulkuväylänä. Voimakas vulkaaninen toiminta ja merenpinnan lasku sulkivat tämän väylän, mikä johti saatavilla olevan saaliin vähenemiseen ja merivirtojen dramaattiseen muuttumiseen. Oikealla ylhäällä on Keski-Amerikka sellaisena kuin tunnemme sen nykyään, ja siellä on lämpimän veden merivirtaukset.
Pienempien hampaiden tutkiminen osoittaa, että C. megalodonit olivat kasvualueilla jopa 2-3 metrin pituisia. Tämä ei kuitenkaan tarkoita sitä, että ne olisivat olleet näin isoja syntyessään, vaan ainoastaan sitä, että hampaat olivat peräisin nuorilta yksilöiltä, jotka olivat aktiivisia tällä alueella.Kun nuoret C. megalodonit kasvoivat vanhemmiksi, ne kasvaisivat yhä suuremmiksi, ja näin ollen niiden olisi lopulta lähdettävä matalikoista avomerielämään. Tällöin alkaisi niiden elämän toinen vaihe, jossa niiden olisi erikoistuttava hyökkäämään suurten valtamerieläinten kimppuun.
Saaliseläimet – Mitä megalodonit söivät?
On välittömästi selvitettävä, että C. megalodondon huomasi dinosaurukset. Tämä on myytti, jota levitetään populaarikulttuurissa, erityisesti elokuvissa ja romaaneissa, joissa C. megalodonin ”pahis” rakennetaan dramaattisemmaksi. C. megalodondin esiintyminen fossiilirekisterissä alkaa vasta myöhäisligoseenissä, noin 36 miljoonaa vuotta sen jälkeen, kun dinosaurukset kuolivat sukupuuttoon liitukauden lopussa, mikä tekee C. megalodondin esiintymisen mahdottomaksi. megalodon olisi voinut syödä dinosauruksia (hai, joka todella olisi voinut syödä dinosauruksia ja suuria merellisiä matelijoita, löytyy osoitteestaCretoxyrhina).
C. megalodonin suosima ravinto näyttää olleen valaita ja erityisesti pieniä ja keskikokoisia valaita. On myös todisteita siitä, että C. megalodon on voinut syödä dinosauruksia ja suuria merellisiä matelijoita.megalodon hyökkäsi ja söi suuria merikilpikonnia, jotka olivat oletettavasti liian hitaita paetakseen, vaikka niiden kuori ei suojannut C. megalodonin kolossaaliselta puremavoimalta. Se, mitä C. megalodon metsästi, riippui kuitenkin yksilön iästä, sillä pienemmät C. megalodonit metsästivät eläimiä, kuten vuohia, ja isommat, vanhemmat C. megalodonit metsästivät isompia valaita.
C. megalodonin hyökkäysstrategia ei ollut riittävä. Joidenkin valaiden nikamissa on puristumisvaurioita, joiden on tulkittu johtuneen äkillisestä ja massiivisesta alhaalta tulevasta iskusta. Tämän perusteella on voitu rekonstruoida skenaario, jossa C. megalodon lähestyi valasta alhaalta päin välttääkseen kohteensa näkemisen.Kun se oli valmistautunut iskuun, C. megalodon käytti voimakkaita lihaksiaan ponnahtaakseen pintaan suurella nopeudella ja iskeytyäkseen valaan päälle alapuolelta. Jos valas ei olisi päätynyt C. megalodonin leukoihin, se olisi hyvin todennäköisesti tainnutettu törmäyksestä, jolloin C. megalodonilla olisi ollut aikaa tappavaan puremaan.Tiedetään kuitenkin ainakin yksi fossiilinen nikama, joka osoittaa, että se oli joutunut tämäntyyppisen hyökkäyksen kohteeksi, mutta onnistui silti parantumaan. Tämä osoittaa, että tässä tapauksessa onnekas eläin ei vain selvinnyt hyökkäyksestä, vaan se myös eli niin kauan, että vammat ehtivät parantua.

Erityishuomautus* – Yllä olevan sekvenssin tarkoituksena on havainnollistaa erästä mahdollista tapaa, jolla C.-megalodon-hait metsästivät saalista. Tarkoituksena ei ole viitata siihen, että tämä olisi ollut ainoa tapa, jolla C. megalodon-hait saalistivat.
C. megalodon-haiden saaliseläimistä ilmeisesti peräisin olevien fossiilien tutkiminen osoittaa, että C. megalodon-hait kohdistivat saaliinsa itse asiassa luisiin alueisiin, kuten niskahäkkiin. Tällöin C. megalodon-haitilla oli kaksi asiaa edukseen: erittäin vankat hampaat, jotka eivät murtuneet helposti, ja murskaava puraisuvoima, jonka avulla voitiin helposti murskata luita, jotka puolestaan aiheuttivat laajamittaisia vammoja sisäelimissä, joita niiden oli tarkoitus suojella.Lisätukea tälle menetelmälle antavat hampaiden puristusmurtumat, joissa hampaat ovat tylpistyneet, mikä viittaa voimakkaisiin iskuihin luun kaltaiseen kovaan aineeseen.
Haukatessaan isompia valaita, jotka olivat todennäköisesti liian isoja puremaan kylkiluiden kaltaiseen alueeseen, C. megalodon muutti toimintatapaansa. Kriittisten elinten kimppuun hyökkäämisen sijaan C. megalodon hyökkäsi hännän kimppuun yrittäessään liikuntakyvyttömäksi saalistaan.Tämä on erittäin älykäs strategia, sillä vaikka hait uivat lähes jatkuvasti eteenpäin, jotta ne voivat hengittää, ne pystyvät ylläpitämään erittäin nopeaa takaa-ajonopeutta vain lyhyen aikaa. Tämä johtuu siitä, että haiden valkoiset lihakset (noin 90 prosenttia koko lihasmassasta), joita käytetään äkillisiin nopeuspyrähdyksiin, väsyvät hyvin nopeasti, kun taas haiden punaiset lihakset (noin 10 prosenttia koko lihaksistosta) ovat teholtaan heikompia mutta kestävyydeltään uskomattoman kestäviä, minkä vuoksi punaisia lihaksia käytetään tavanomaiseen matkantekoon.Rampauttamalla suuren valaan C. megalodon voisi rauhassa syödä sen sijaan, että se ponnistelisi liikaa.
Jotkut ihmiset ovat väittäneet, että C. megalodon olisi ollut yksinkertaisesti liian suuri metsästääkseen ja että se olisi voinut olla vain haaskaeläin. Kun otetaan huomioon ylivoimainen fossiilinen todistusaineisto, joka osoittaa vammoja, ei vain hampaiden jättämiä jälkiä, monille suurille valaaneläimille, tällaista väitettä ei pidetä vain epätodennäköisenä, vaan melkeinpä jopa mahdottomana.Vaikka useimmat hait ja lihansyöjäeläimet yleensä tarttuvat tilaisuuteen syödä raadosta, se ei tee niistä yksinomaan haaskalintuja. Myös merieläimet, jotka elävät pelkällä haaskalintujen syönnillä, ovat yleensä pohjasyöjiä, jotka odottavat kuolleiden eläinten vajoamista merenpohjaan. C.megalodonin koon arvioiminen on myös synnyttänyt arvioita siitä, kuinka paljon ravintoa se tarvitsisi elääkseen.Määrät vaihtelevat suuresti, mutta ne vaihtelevat 600-1200 kilon välillä joka päivä. Tämä on valtava määrä ruokaa raadonsyöjälle, sillä raadonsyöjillä on tapana sopeutua siihen, että ne tarvitsevat hyvin vähän energiaa, koska ne eivät tiedä, milloin tai mistä seuraava ateria on tulossa.Kun otetaan huomioon kaikki fossiiliset todisteet, biometriset mallit ja hain elintapoja ja biologiaa koskeva tietämys, tulokseksi saadaan, että raadonsyönti on vähiten todennäköinen tapa selviytyä hengissä C.megalodonilla.

Katoaminen – Miksi megalodon katosi?
C.megalodondis katoaa fossiiliaineistosta lähellä pleistoseenin ensimmäisen vaiheen (gelasian) loppua1,8 miljoonaa vuotta sitten. Tätä katoamista leimaa C.megalodondiksen tasainen väheneminen. Vaikka on olemassa muutamia teorioita siitä, miksi C. megalodon katosi, näyttää todennäköisimmältä, että C. megalodonin katoaminen johtui useista muuttuvista tapahtumista eikä vain yhdestä asiasta.
C. megalodonin sukupuuttoon kuolemisen laukaiseva tapahtuma näyttää olleen maailmanlaajuinen jäähtyminen. Aluksi, jos C. megalodonilla oli lämminverinen aineenvaihdunta jättilämmön ansiosta, se tarvitsisi enemmän kaloreita kuin samankokoinen täysin kylmäverinen olento. Mitä kylmempi vesi on, sitä suurempi ero on, mikä tarkoittaa, että C. megalodon olisi tarvinnut vieläkin enemmän ravintoa selviytyäkseen lämpötilan laskusta. Lisäksi jättilämpö ei edelleenkään korvaa todellista lämminveristä aineenvaihduntaa, ja viileämmissä vesissä elävä hai olisi silti mahdollisesti ollut vaisumpi kuin lämpimissä vesissä, mikä vaikeuttaisi entisestään sen metsästysvalmiuksia.
Kylmemmän maapallon lämpötilan vaikutuksesta suuret vesimassat alkoivat jähmettyä jääksi, mistä ovat osoituksena valtavat jääpeitteet pohjoisella pallonpuoliskolla.Jään lisääntyminen merkitsi sitä, että merenpinta laski, ja dramaattisin seuraus tästä oli Panaman kannaksen syntyminen, mitä edesauttoi myös se, että alueella jatkuva tulivuoritoiminta synnytti uusia maamuodostumia.Tämä loi pohjimmiltaan maasillan Pohjois- ja Etelä-Amerikan välille sekä erotti Tyynenmeren ja Atlantin valtameren toisistaan tässä vaiheessa.
Panaman kannaksen syntymisen välittömänä seurauksena sulkeutui Keski-Amerikan meriväylä, jota näytti käytetyn valaiden tärkeimpänä vaellusreittinä, mistä on osoituksena valasfossiilien suuri määrä.Tämä ajoittuu samaan aikaan valaslajien yleisen vähenemisen kanssa, sillä alle puolet pleistoseenin valaslajeista on säilynyt elossa nykyaikaan asti. Nykyään on vain kuusi valassukua, kun taas mioseenin aikana niitä oli yli 20. Jäljelle jääneet valaat vaelsivat edelleen, mutta ne näyttivät suosivan polaarialueita, luultavasti siksi, että siellä oli enemmän selkärangattomia eläimiä, joiden syömiseen paaluvalaat ovat sopeutuneet.Myöskään hammasvalaat eivät näytä olleen käyttökelpoinen vaihtoehto, sillä myös niiden määrä väheni dramaattisesti, ja siittiövalas on ainoa suuri hammasvalas, joka on säilynyt hengissä nykypäivään asti. Kun C. megalodon rajoittui lämpimämpiin merivesiin, sillä ei enää ollut ympärivuotista ja jatkuvaa pääsyä ravintoon, jonka tappamiseen se oli sopeutunut parhaiten.
C. megalodonin valtava koko oli epäilemättä haitannut sitä näinä aikoina, sillä ainoat muut saatavilla olevat saaliseläimet olivat pienempiä ja nopeampia, ja vaikka niitä olisi saatu, ne eivät tarjonneet samanlaista ravintoa kuin suuremmat valaat.Kannibalismia on myös ehdotettu C. megalodonin mahdolliseksi selviytymisstrategiaksi, mutta se toimisi vain niin kauan kuin olisi muita C. megalodoneja syötäväksi. Jos näin todella tapahtuisi, kannibalismi vain harventaisi C. megalodonin määrää entisestään, mikä puolestaan rajoittaisi lisääntymiskykyistä määrää.
Tähän liittyy merenpinnan korkeuden muuttumisen mahdollisesti aiheuttama taimitarha-alueiden menetys.Itse asiassa Panaman kannaksen luominen näyttää poistaneen yhden tällaisen kasvualueen, mistä on osoituksena suuri määrä nuorten C. megalodonien hampaita tältä alueelta.Toinen epäilty kasvualue oli Maryland, joka sijaitsi niin kaukana pohjoisessa, että vedet saattoivat muuttua liian kylmiksi C. megalodonien elinympäristöksi. Kasvualueiden häviäminen merkitsee sitä, että C. megalodonin poikaset olisivat olleet alttiimpia petoeläimille, kenties jopa toisille C. megalodonille, jotka yrittivät selviytyä.
Viimeiseen teoriaan liittyy uusien petoeläinten ilmaantuminen, ja siinä viitataan erityisesti petoeläimiin kuuluvien delfiinien evoluutioon, joita nykyään edustaa Orca, joka tunnetaan myös nimellä miekkavalas. C. megalodonin määrän vähentyessä delfiinien määrä kasvoi.On kuitenkin vaikea sanoa, oliko näiden uusien petoeläinten nousulla osuutta C. megalodonin taantumiseen, sillä yhtä hyvin C. megalodonin taantuminen saattoi antaa uusille petoeläimille tilaa menestyä.On olemassa fossiilisia todisteita, jotka osoittavat, että C. megalodonin ja C. megalodonin välillä on petoeläimen ja saaliin välistä vuorovaikutusta. megalodonin ja delfiniidien välillä, mistä kertovat C.megalodonin hampaanjäljet delfiniidien luissa.
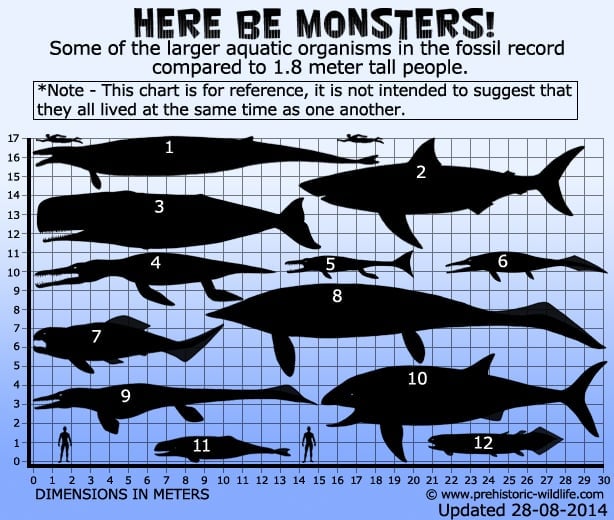
1 – Basilosaurus(valas), 2 – C.megalodon – alempi keskiarvo-arvio (hai), 3 – Livyatanmelvillei – alempi arvio (valas), 4 – Pliosaurusfunkei, a.k.a Predator X (pliosaurus), 5 -Plesiosuchus(talattosuchian), 6 – Thalattoarchon(iktyosaurus), 7 – Dunkleosteus(arthrodire placoderm), 8 – Shastasaurus(iktyosaurus),9 – Tylosaurus(mosasaurus), 10 – Leedsichthys- ylempi estimaatti(kalat)), 11 – Brygmophyseter(valas), 12- Rhizodus(lohko-uimapyrstöinen kalalaji).
Viimeiset eloonjääneet?
Joidenkin mielestä C. megalodon selviytyi pleistoseenikaudelta ja ui valtamerissä vielä holoseenikaudella. Heidän todisteensa tästä väitteestä on HMS Challenger -aluksen miehistön vuonna 1872 löytämä C. megalodonin osittainen hammas, jonka vuonna 1959 tehdyissä testeissä sen uskottiin olevan vain 10 000 vuotta vanha.Tässä testissä mitattiin kuitenkin fossiilin mangaanidioksidipitoisuuksia, ja tätä menetelmää pidetään nykyään virheellisenä, koska mangaanidioksidipitoisuudet voivat vaihdella eri fossiileissa, jopa samalta aikakaudelta peräisin olevissa fossiileissa. Kun hammasta tutkittiin myöhemmillä radiohiiliajoitusmenetelmillä, hampaan typpipitoisuus osoittautui liian alhaiseksi, jotta sitä olisi voitu testata, joten hammasta ei ole sittemmin voitu testata, ja aiemmat arviot C. megalodonin sukupuuttoon kuolemisesta ulos pleistoseenikauden alkupuoliskon aikana ovat edelleen voimassa.

Luokittelu – Liittyykö megalodon valkohaiin?
Ehkä suurin kiistanaihe C. megalodonista on se, liittyykö se todella nykyisin tuntemaamme valkohaiin. Vertailuja C. megalodonin ja valkohain välillä on tehty mm. megalodonin ja valkohain väliset vertailut perustuvat useimmiten siihen periaatteeseen, että C. megalodon oli suurin hai, ja valkohai on suurin hai, jonka tunnemme nykyään. Myös näiden kahden hain hampaita pidetään usein suunnilleen samanlaisina.C. megalodonin kannattajat viittaavat siihen, että hampaiden samankaltaisuus johtuu siitä, että sekä C. megalodon että valkohai polveutuvat Palaeocarcharodonorientaliksesta.
Ongelma C. megalodonin ja valkohain vertailussa on se, että C. megalodonin ja valkohain hampaiden samankaltaisuus johtuu siitä, että sekä C. megalodon että valkohai polveutuvat Palaeocarcharodonorientaliksesta. megalodonin ja valkohaiden vertailu samankaltaisten hampaiden perusteella johtuu siitä, että ainoat yhtäläisyydet ovat siinä, että molemmilla hailla on kolmionmuotoiset ja hammastetut hampaat. Tämän lisäksi valkohaiden hampaat ovat paljon ohuemmat kuin C. megalodonilla, koska ne ovat paljon ohuemmat kuin C. megalodonilla. Vaikka C. megalodonin ja valkohaiden hampaat ovat paljon ohuemmat.megalodonilla uskotaan olevan kaiken kaikkiaan samanlainen hammasrakenne kuin valkohailla, C. megalodonin kolmas etuhammas (kolmas etuhammas yläleuan keskeltä) on erilainen siinä mielessä, että se osoittaa alaspäin kahden ensimmäisen hampaan tapaan, toisin kuin valkohailla. C. megalodonin etuhampaissa on myös tyypillinen ”arpi”, joka on muodoltaan sakaran muotoinen ja joka sijaitsee hampaan kruunun ja juuren välissä, mikä puuttuu valkohaista.
Vaihtoehto C. megalodonin sijoittamiselle Carcharodonshaiden sukuun olisi sen sijoittaminen vanhempaan Carcharoclesgenus-sukuun. Tärkein peruste tälle sijoittamiselle on se, että erään toisen suuren muinaisen hain nimeltäCarcharoclesauriculatus uskotaan itse asiassa olevan C. megalodonin esi-isä. Lähes 12 senttimetrin pituisten hampaidensa ansiosta Carcharoclesauriculatus oli suuri, mutta noin kolmanneksen pienempi kuin C. megalodon, jos se suhteutetaan 18-senttiseen C. megalodoniin. Siitä huolimatta on täysin mahdollista, että Carcharoclesauriculatus on kasvanut suuremmaksi, jolloin on syntynyt C. megalodon, koska petoeläimillä on kaikissa ympäristöissä taipumus kasvaa yhä suuremmiksi, kunnes niiden ympäristö ei enää kestä kasvua.
Tällainen C. megalodonin sijoittuminen Carcharoclesin sisälle päättäisi itse asiassa siirtymävaiheen, jossa hait menettivät hampaidensa sivuhampaita.Tämä siirtymä alkaa hampaiden sivuhampaista, joita on selvästi Otodusobliquuksessa, Carcharoclesauriculatusin vähentyneistä hampaiden sivuhampaista ja päättyy hampaiden puuttumiseen C. megalodonissa.Näissä hampaissa on myös chevroninmuotoisia arpia, joissa kruunu yhtyy juureen, mikä puuttuu suurista valkoisista hampaista.
Muiden teorioiden mukaan C. megalodon oli suuren valkohain esi-isä, ja ajan mittaan hai yksinkertaisesti pieneni. Suurin ongelma tässä ajattelussa on kuitenkin se, että suuri valkohai ui valtameressä jo kauan ennen C. megalodonia. megalodon kuoli sukupuuttoon, ja valkohain fossiilihampaat ilmestyivät jo 16 miljoonaa vuotta sitten, yli 14 miljoonaa vuotta ennen kuin C. megalodon kuoli sukupuuttoon. Teorian kannattajat väittävät edelleen, että valkohai on voinut kehittyä pienemmästä C. megalodonin lajista. Ne, jotka tuntevat sapelihammaskissa Smilodonin, saattavat tietää, että samasta suvusta oli olemassa kolme aivan erilaista lajia, jotka eivät ainoastaan näytä polveutuvan samasta esi-isästä, vaan myös toimineet jonkin aikaa yhdessä samalla ajanjaksolla.Ei ole kovin kaukaa haettua, että vastaavaa voisi tapahtua muissakin eläinryhmissä. Silti ei näytä olevan mitään siirtymävaihetta, joka osoittaisi, että C. megalodonin hampaat eivät vain pienentyisi, vaan ne muuttuisivat tasaisesti suuren valkoisen muodon mukaisiksi.Jos ne todella ovat sukua, on todennäköisempää, että suurella valkohampaalla on yhteinen välitön esi-isä C. megalodonin kanssa.
Vuonna 2012 teoria, jonka mukaan C. megalodon ja Carcharodoncarcharias eivät ole sukua toisilleen, sai hieman lisää tukea, kun kuvailtiin uusi Carcharodonin laji, Carcharodonhubbelli, joka tunnetaan myös nimellä Hubbellin valkohai. Jotkut ovat tulkinneet Carcharodon hubbellin hampaiden olevan muodoltaan siirtymävaiheen hampaita, jotka yhdistävät Carcharodon carcharias -lajin Isurusgenus-sukuun, johon makohait kuuluvat. Koska useimmat tutkijat eivät katso, että makohain ja C. megalodonin kaltaisten valkohaiden välillä on suora yhteys, tämä saattaa viitata siihen, että Carcharodon carcharias on todellakin erillinen C. megalodonista ja että C. megalodon olisi näin ollen sijoitettava Carcharoclesgenus-sukuun.
Mahdolliset yhtäläisyydet makohain ja valkohain yleisessä morfologiassa ovat todennäköisesti seurausta pikemminkin evoluutiosta kuin geneettisestä jalostuksesta.Tätä perusruumiin muotoa kutsutaan fuusionmuotoiseksi tai väljemmin ”torpedonmuotoiseksi”, ja se perustuu terävään etuosaan, joka nousee leveään keskikohtaan, ennen kuin se kapenee toiseen takimmaiseen pisteeseen. Tämä muoto on toistunut luonnossa lukemattomia kertoja, eikä se todellakaan ole ainutlaatuinen vain hailla, sillä se on yksinkertaisesti tehokkain muoto vedenalaisessa vesiliikenteessä. Joidenkin mielestä megalodonhain on myös katsottu kuuluvan joko Procarcharodon- tai Otodus-sukuun. Tämä ajattelu perustuu tutkimuksiin, joiden mukaan Otodus-tyypin hampaista on havaittavissa siirtymä Otodus-suvun hampaista myöhemmin eläneen megalodonlajin hampaisiin. Jälleen kuitenkin tutkijoiden väliset erot vaihtelevat suuresti sen suhteen, mihin sukuun megalodonlajit kuuluvat.
Lisälukemista
– Recherches sur les poissons fossiles/par Louis Agassiz – Neuchatel:Petitpierre. s. 41. – Louis Agassiz – 1833-1843.
– Suuren valkohain (Carcharodon) koko – Science Magazine 181(4095): 169-170 – John Randall – 1973.
– Carcharodon megalodon from the Upper Miocene ofDenmark, withcomments on elasmobranch tooth enameloid: coronoi’n – Bulletin of theGeological Society of Denmark (Kööpenhamina: Geologisk Museum) 32: 1-32.- Svend Erik Bendix-Almgreen – 1983.
– Catalogue of Cuban fossil Elasmobranchii (Paleocene to Pliocene) andpaleogeographic implications of their Lower to Middle Mioceneourrenceoccurrence – Boletín de la Sociedad Jamaicana de Geología (Cuba) 31:7-21 – M. Iturralde-Vinent, G. Hubbel & R. Rojas – 1996.
– The Megatooth shark, Carcharodon megalodon: Roughtoothed, hugetoothed – Mundo Marino Revista Internacional de Vida (non-refereed)(Marina) 5: 6-11. – J. C. Bruner – 1997.
– Fossil sharks from Jamaica – Bulletin of the Mizunami Fossil Museum.pp. 211-215. – Stephen Donovan & Gunter Gavin – 2001.
– An associated specimen of Carcharodon angustidens(Chondrichthyes,Lamnidae) from the Late Oligocene of New Zealand, with comments onCarcharodon interrelationships – Journal ofVertebrate Paleontology 21(4): 730-739. – M. D. Gottfried & R. E. Fordyce – 2001.
– The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes:Lamnidae) – Journalof Fossil Research (Japan) 35 (2): 28-33. – Kenshu Shimada – 2002.
– New Record of the Lamnid Shark Carcharodon megalodonf from the MiddleMiocene of Puerto Rico – Caribbean Journal of Science 39: 223-227. -Angel M. Nieves-Rivera, Maria Ruizyantin & Michael D. Gottfried- 2003.
– The Miocene Climatic Optimum: evidence from ectothermic vertebratesof Central Europe – Palaeogeography, Palaeoclimatology, Palaeoecology195 (3-4): 389-401 – M. Böhme – 2003.
– Carcharocles megalodonin (Lamniformes:Otodontidae) ikä: katsaus stratigrafisiin tietoihin – The Palaeontological Society of Japan(PSJ) (Japan) 75 (75): 7-15. – Hebe Hideo, Goto Mastatoshi &Kaneko Naotomo – 2004.
– Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from theVenezuela Neogene: Their Role in the Caribbean, Shallow-water FishAssemblage – Caribbean Journal of Science 40 (3): 362-368. – O.Aguilera & E. R. D. Aguilera – 2004.
– Tracing the ancestry of the Great White Shark – Journal of VertebratePaleontology 26 (4): 806-814 – K. G. Nyberg, C. N. Ciampaglio &G. A. Wray – 2006.
– Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms – The Journal of Geology (USA: The Universityof Chicago) 104 (2): 143-162. – Warren D. Allmon, Steven D. Emslie,Douglas S. Jones & Gary S. Morgan – 2006.
– Three-dimensional computer analysis of white shark leukamekaniikan kolmiulotteinen tietokoneanalyysi: kuinka kovaa valkohai voi purra? – Journal of Zoology 276 (4): 336-342. -S. Wroe, D. R. Huber, M. Lowry, C. McHenry, K. Moreno, P. Clausen, T.L. Ferrara, E. Cunningham, M. N. Dean & A. P. Summers – 2008.
– Miocene sharks in the Kendeace and Grand Bay formations of Carriacou,The Grenadines, Lesser Antilles – Caribbean Journal of Science. 44 (3)pp. 279-286. – Roger Portell, Gordon Hubell, Stephen Donovan, JeremyGreen, David Harper & Ron Pickerill – 2008.
– Giant-toothed white sharks and cetacean trophic interaction from thePliocene Caribbean Paraguaná Formation – Paläontologische Zeitschrift(Springer Berlin) 82 (2): 204-208. – Orangel A. Augilera, Luis García *Mario A. Cozzuol – 2008.
– Ancient Nursery Area for the Extinct Giant Shark Megalodonfrom theMiocene of Panama – PLoS ONE (Panama: PLoS.org) 5 (5): e10552 -Catalina Pimiento, Dana J. Ehret, Bruce J. McFadden & GordonHubbell – 2010.
– The Great White Shark Carcharodon carcharias(Linne, 1758) in thePliocene of Portugal and its Early Distribution in Eastern Atlantic -Revista Española de Paleontología (Portugali) 25 (1): 1-6. – MiguelTelle Antunes, Ausenda Cáceres Balbino – 2010.
– Patterns and ecosystem consequences of shark declines in the ocean -Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055-1071. -Francesco Ferretti, Boris Worm, Gregory L. Britten, Michael J. Heithaus& Heike K. Lotze – 2010.
– Origin of the white shark Carcharodon(Lamniformes: Lamnidae)based on recalibration of the upper Neogene Pisco Formation of Peru- Palaeontology 55(6):1139-1153 – D. J.Ehret, B.J. MacFadden, D. S. Jones, T. J. DeVries, D. A.Foster & R. Salas-Gismondi – 2012.
– Valkoisten ja megahampaiden haiden evoluutio ja todistusaineisto hylkeisiin, sireeneihin ja valaisiin kohdistuvasta varhaisesta petoeläimistä – Luonnontieteellinen tiede (TšekkiläinenRepublikaani) 5 (11): 1203-1218. – C. G. Diedrich – 2013.
– Sharks and Rays (Chondrichthyes, Elasmobranchii) from the LateMiocene Gatun Formation of Panama – Journal of Paleontology 87 (5):755-774 – Catalina Pimiento, Gerardo González-Barba, Dana J. Ehret,Austin J. W. Hendy, Bruce J. MacFadden & Carlos Jaramillo -2013.
– When Did Carcharocles megalodon Become Extinct? ANew Analysis of theFossil Record. – PLOS ONE. 9 (10): e111086. – C. Pimiento & C.F. Clements – 2014.
– Body-size trends of the extinct giant shark Carcharoclesmegalodon: adeep-time perspective on marine apex predators. – Paleobiology. 41 (3):479-490. – C. Pimiento & M. A. Balk – 2015.
– Record of Carcharocles megalodon in the EasternGuadalquivir Basin(Upper Miocene, South Spain). Estudios Geológicos. 71 (2): e032. – M.Reolid & J. M. Molina – 2015.
– Geographical distribution patterns of Carcharocles megalodonovertime reveal clues about extinction mechanisms. – Journal ofBiogeography. 43 (8): 1645-1655. – C. Pimiento, B. J. MacFadden, C. F.Clements, S. Varela, C. Jaramillo, J. Velez-Juarbe & B. R.Silliman – 2016.
– The size of the megatooth shark, Otodus megalodon(Lamniformes:Otodontidae), revisited. – Historical Biology: 1-8. – Kenshu Shimada -2019.
– The Early Pliocene extinction of the mega-toothed shark Otodusmegalodon: a view from the Eastern North Pacific. – PeerJ. 7:e6088. -R. W. Boessenecker, D. J. Ehret, D. J. Long, M. Churchill, E. Martin& S. J. Boessenecker – 2019.
– The transition between Carcharocles chubutensisand Carcharoclesmegalodon (Otodontidae, Chondrichthyes): lateral cusplet lossthrought time. – Journal of Vertebrate Paleontology. 38 (6): e1546732. – V. J.Perez, S. J. Godfrey, B. W. Kent, R. E. Weems & J. R. Nance -2019.
– Body dimensions of the extinct giant shark Otodus megalodon:a 2Dreconstruction. – Scientific Reports. 10 (14596): 14596. – J. A.Cooper, C. Pimiento, H. G. Ferrón & M. J. Benton – 2020.
—————————————————————————-
Sattumanvaraiset suosikit
 |
 |
|
 |
 |
Tietosuoja& Evästekäytäntö