 L’Anthocerotophyta est le phylum le moins diversifié des bryophytes, cependant sa distribution est étendue, l’Antarctique étant le seul continent où on ne les trouve pas. La plupart des espèces de cet embranchement poussent généralement sur des sols humides dans des zones ombragées, mais certaines poussent dans des sites exposés, tandis que d’autres sont épiphytes. Les calicots sont considérés comme des pionniers écologiques, car ils poussent souvent dans des zones où il y a peu ou pas de concurrence, comme sur les sols minéraux. Bien qu’ils puissent ressembler superficiellement à une hépatique thalloïde, ils ont des caractéristiques gamétophytiques et sporophytiques qui les séparent des autres bryophytes vus jusqu’à présent.
L’Anthocerotophyta est le phylum le moins diversifié des bryophytes, cependant sa distribution est étendue, l’Antarctique étant le seul continent où on ne les trouve pas. La plupart des espèces de cet embranchement poussent généralement sur des sols humides dans des zones ombragées, mais certaines poussent dans des sites exposés, tandis que d’autres sont épiphytes. Les calicots sont considérés comme des pionniers écologiques, car ils poussent souvent dans des zones où il y a peu ou pas de concurrence, comme sur les sols minéraux. Bien qu’ils puissent ressembler superficiellement à une hépatique thalloïde, ils ont des caractéristiques gamétophytiques et sporophytiques qui les séparent des autres bryophytes vus jusqu’à présent.
Toutes les espèces ont un gamétophyte thalloïde qui est aplati et peut se présenter sous forme de rosette ou de ruban. 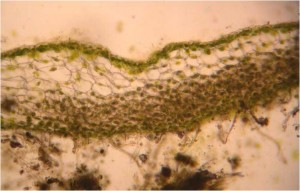 Dans la plupart des genres, le thalle est multistrateux au centre et s’amincit plus près des marges. Les cellules de la surface supérieure du thalle sont généralement chlorophylliennes, tandis que celles qui se trouvent dans les couches plus profondes ne contiennent pas de chloroplastes. La plupart des genres de ce phylum possèdent un seul grand chloroplaste par photosynthèse ainsi qu’un pyrénoïde associé au plaste. Les cormorans regroupent la Rubisco dans le pyrénoïde, ce qui est un trait que l’on observe également chez les algues. Ces caractéristiques, ainsi que l’absence de corps gras, aident à distinguer ces espèces de celles du Marchantiophyta. La surface du thalle est généralement cutinée, mais cette couche cuticulaire n’empêche pas les échanges gazeux de se produire. Les thalles sont attachés aux substrats par des rhizoïdes lisses, à parois minces, non ramifiés et unicellulaires.
Dans la plupart des genres, le thalle est multistrateux au centre et s’amincit plus près des marges. Les cellules de la surface supérieure du thalle sont généralement chlorophylliennes, tandis que celles qui se trouvent dans les couches plus profondes ne contiennent pas de chloroplastes. La plupart des genres de ce phylum possèdent un seul grand chloroplaste par photosynthèse ainsi qu’un pyrénoïde associé au plaste. Les cormorans regroupent la Rubisco dans le pyrénoïde, ce qui est un trait que l’on observe également chez les algues. Ces caractéristiques, ainsi que l’absence de corps gras, aident à distinguer ces espèces de celles du Marchantiophyta. La surface du thalle est généralement cutinée, mais cette couche cuticulaire n’empêche pas les échanges gazeux de se produire. Les thalles sont attachés aux substrats par des rhizoïdes lisses, à parois minces, non ramifiés et unicellulaires.
Alors que le thalle est principalement composé de cellules parenchymes et présente peu de différenciation tissulaire, 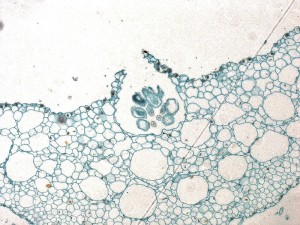 il y a souvent des cavités intercellulaires, qui sont remplies de mucilage, sur la surface ventrale. Ces chambres à mucilage, comme on peut le voir sur cette image, s’ouvrent sur l’environnement grâce à la présence de pores ressemblant à des stomates et sont fréquemment envahies par de petites colonies d’algues bleu-vert appelées Nostoc.
il y a souvent des cavités intercellulaires, qui sont remplies de mucilage, sur la surface ventrale. Ces chambres à mucilage, comme on peut le voir sur cette image, s’ouvrent sur l’environnement grâce à la présence de pores ressemblant à des stomates et sont fréquemment envahies par de petites colonies d’algues bleu-vert appelées Nostoc.
Les Nostoc sont des cyanobactéries filamenteuses qui peuvent facilement fixer l’azote. Elles sont capables de former des relations symbiotiques avec les hornworts dans lesquelles l’algue reçoit des hydrates de carbone et un abri protégé tandis que le hornwort reçoit l’azote fixé.
La plupart des espèces de ce phylum sont bisexuées, ayant à la fois l’organe sexuel mâle et femelle intégré dans la surface supérieure du thalle. Les anthéridies, qui se développent dans des chambres à l’intérieur du thalle, sont issues d’une seule initiale. De plus, cette initiale est capable de donner naissance à plusieurs anthéridies au sein d’une même chambre, comme on le voit sur cette image. 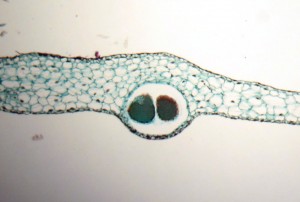 Les archégones, qui sont également enchâssés dans le thalle, sont issus d’une cellule initiale. Alors que seule la partie supérieure du col s’élève de la surface du thalle, la partie inférieure du col ainsi que la cellule-œuf restent sous la surface. Comme les cellules qui entourent l’archégone ne diffèrent pas de celles du thalle, l’organe sexuel n’est pas considéré comme discret.
Les archégones, qui sont également enchâssés dans le thalle, sont issus d’une cellule initiale. Alors que seule la partie supérieure du col s’élève de la surface du thalle, la partie inférieure du col ainsi que la cellule-œuf restent sous la surface. Comme les cellules qui entourent l’archégone ne diffèrent pas de celles du thalle, l’organe sexuel n’est pas considéré comme discret.
Les Anthocerotophyta sont caractérisés par leur sporophyte long, en forme de corne, d’où leur nom de « hornworts ». Bien que le sporophyte reste attaché à son parent, comme chez les autres bryophytes, il diffère significativement en ce qu’il possède une région méristématique à sa base. Cette caractéristique permet au sporophyte d’avoir une croissance indéterminée, lui donnant la capacité de différencier de nouveaux tissus sporogènes tout au long de sa vie. Des stomates similaires à ceux des plantes vasculaires peuvent être trouvés sur le sporangium, cependant, contrairement à ceux des plantes supérieures, ils ne s’ouvrent pas ou ne se ferment pas.
Bien que le sporophyte reste attaché à son parent, comme chez les autres bryophytes, il diffère significativement en ce qu’il possède une région méristématique à sa base. Cette caractéristique permet au sporophyte d’avoir une croissance indéterminée, lui donnant la capacité de différencier de nouveaux tissus sporogènes tout au long de sa vie. Des stomates similaires à ceux des plantes vasculaires peuvent être trouvés sur le sporangium, cependant, contrairement à ceux des plantes supérieures, ils ne s’ouvrent pas ou ne se ferment pas.
A mesure que le sporophyte grandit, il devient non seulement plus long mais les spores à différentes hauteurs dans le sporangium diffèrent en maturité.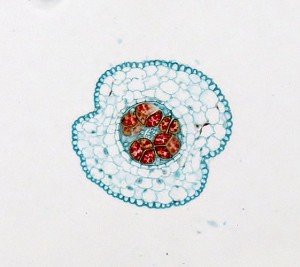 Les spores plus jeunes à la base du sporangium restent typiquement dans un arrangement tétraédrique, tandis que les spores matures près de l’extrémité sont séparées. Le sporophyte est dépourvu de seta mais possède une columelle, et la déhiscence se produit le long d’une ou deux lignes longitudinales, ainsi les spores sont libérées le long des moitiés à mesure qu’elles mûrissent.
Les spores plus jeunes à la base du sporangium restent typiquement dans un arrangement tétraédrique, tandis que les spores matures près de l’extrémité sont séparées. Le sporophyte est dépourvu de seta mais possède une columelle, et la déhiscence se produit le long d’une ou deux lignes longitudinales, ainsi les spores sont libérées le long des moitiés à mesure qu’elles mûrissent.
Non seulement il y a des spores à l’intérieur du sporangium, mais il y a aussi la présence de pseudo-elaters. Ces structures multicellulaires sont filamenteuses 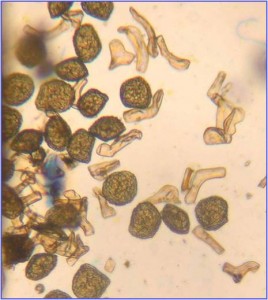 et aident à la dispersion des spores en changeant de forme lorsqu’elles sont sèches. Les parois de leurs cellules peuvent être soit minces, soit épaissies en spirale, soit uniformes. Bien qu’elles ressemblent aux élatères observés chez les hépatiques, la différence dans les modèles de division cellulaire les distingue.
et aident à la dispersion des spores en changeant de forme lorsqu’elles sont sèches. Les parois de leurs cellules peuvent être soit minces, soit épaissies en spirale, soit uniformes. Bien qu’elles ressemblent aux élatères observés chez les hépatiques, la différence dans les modèles de division cellulaire les distingue.
Bien que la reproduction asexuée soit rare chez les hornworts, quelques espèces produisent des gemmes marginales tandis que d’autres sont capables de produire des tubercules pérennes, capables de tolérer une certaine dessiccation. Un moyen simple et efficace de reproduction asexuée consiste à faire mourir les parties les plus anciennes du thalle, laissant les parties plus jeunes et déconnectées poursuivre leur croissance.
Voici quelques exemples d’espèces que l’on trouve dans ce phylum :
Anthoceros spp
Dendroceros spp
Phaeoceros spp
.