Les eucaryotes ont deux grands types de division cellulaire : la mitose et la méiose. La mitose est utilisée pour produire de nouvelles cellules corporelles pour la croissance et la guérison, tandis que la méiose est utilisée pour produire des cellules sexuelles (œufs et spermatozoïdes). La méiose sera abordée dans un chapitre ultérieur.
Le cycle cellulaire est une série ordonnée d’événements impliquant la croissance et la division cellulaire qui produit deux nouvelles cellules filles via la mitose. La durée du cycle cellulaire est très variable, même au sein des cellules d’un organisme individuel. Chez l’homme, la fréquence de renouvellement des cellules va de quelques heures au début du développement embryonnaire à une moyenne de deux à cinq jours pour les cellules épithéliales, ou à une vie humaine entière passée sans division dans les cellules spécialisées telles que les neurones corticaux ou les cellules du muscle cardiaque. Le temps qu’une cellule passe dans chaque phase du cycle cellulaire varie également. Lorsque des cellules de mammifères à division rapide sont cultivées (en dehors du corps dans des conditions de croissance optimales), la durée du cycle est d’environ 24 heures. La chronologie des événements du cycle cellulaire est contrôlée par des mécanismes à la fois internes et externes à la cellule.
Les cellules sur le chemin de la division cellulaire passent par une série d’étapes de croissance, de réplication de l’ADN et de division précisément chronométrées et soigneusement régulées qui produisent deux cellules génétiquement identiques. Le cycle cellulaire comporte deux phases principales : l’interphase et la phase mitotique (figure 1). Pendant l’interphase, la cellule croît et l’ADN est répliqué. Pendant la phase mitotique, l’ADN répliqué et le contenu cytoplasmique sont séparés et la cellule se divise.
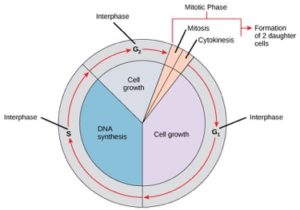
Pendant l’interphase, la cellule subit des processus normaux tout en se préparant à la division cellulaire. Pour qu’une cellule passe de l’interphase à la phase mitotique, de nombreuses conditions internes et externes doivent être réunies. Les trois stades de l’interphase sont appelés G1, S et G2.
Phase G1 (premier écart)
Le premier stade de l’interphase est appelé phase G1 (premier écart) car, d’un aspect microscopique, peu de changements sont visibles. Cependant, pendant la phase G1, la cellule est assez active au niveau biochimique. La cellule accumule les éléments constitutifs de l’ADN chromosomique et des protéines associées, ainsi que des réserves énergétiques suffisantes pour mener à bien la tâche de réplication de chaque chromosome dans le noyau.
Phase S (synthèse de l’ADN)
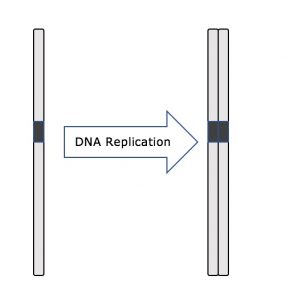
Pendant toute l’interphase, l’ADN nucléaire reste dans une configuration de chromatine semi-condensée. Dans la phase S, la réplication de l’ADN peut se dérouler selon les mécanismes qui aboutissent à la formation de paires identiques de molécules d’ADN – les chromatides sœurs – qui sont fermement attachées à la région centromérique (figure 2).
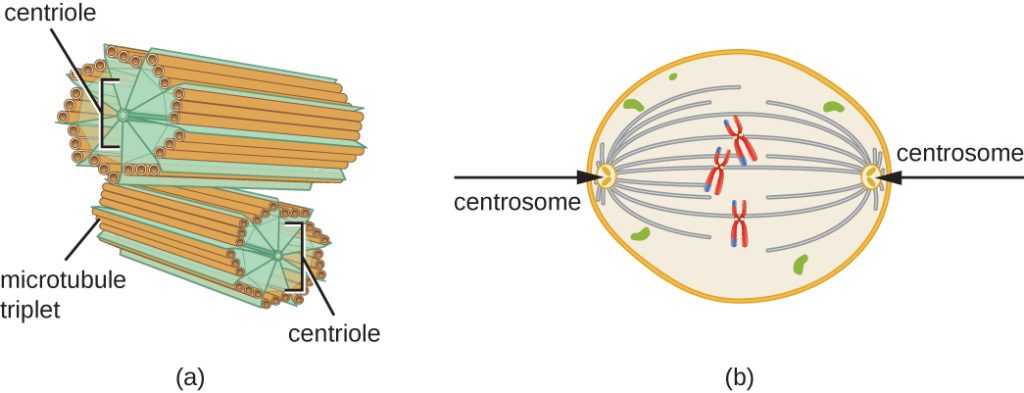
Le centrosome est également dupliqué durant la phase S. Les deux centrosomes vont donner naissance au fuseau mitotique, l’appareil qui orchestre le mouvement des chromosomes pendant la mitose. Au centre de chaque cellule animale, les centrosomes sont associés à une paire d’objets en forme de tige, les centrioles, qui sont perpendiculaires l’un à l’autre. Les centrioles aident à organiser la division cellulaire. Les centrioles ne sont pas présents dans les centrosomes des autres espèces eucaryotes, comme les plantes et la plupart des champignons.
Phase G2 (deuxième écart)
Dans la phase G2, la cellule reconstitue ses réserves énergétiques et synthétise les protéines nécessaires à la manipulation des chromosomes. Certains organites cellulaires sont dupliqués, et le cytosquelette est démantelé pour fournir des ressources pour la phase mitotique. Il peut y avoir une croissance cellulaire supplémentaire pendant la G2. Les derniers préparatifs de la phase mitotique doivent être achevés avant que la cellule puisse entrer dans la première étape de la mitose.
La phase mitotique
Pour faire deux cellules filles, le contenu du noyau et du cytoplasme doit être divisé. La phase mitotique est un processus en plusieurs étapes au cours duquel les chromosomes dupliqués sont alignés, séparés et déplacés vers les pôles opposés de la cellule, puis la cellule est divisée en deux nouvelles cellules filles identiques. La première partie de la phase mitotique, la mitose, est composée de cinq étapes, qui accomplissent la division nucléaire (figure 5). La deuxième partie de la phase mitotique, appelée cytokinèse, est la séparation physique des composants cytoplasmiques en deux cellules filles. Bien que les étapes de la mitose soient similaires pour la plupart des eucaryotes, le processus de la cytokinèse est très différent pour les eucaryotes qui ont des parois cellulaires, comme les cellules végétales.
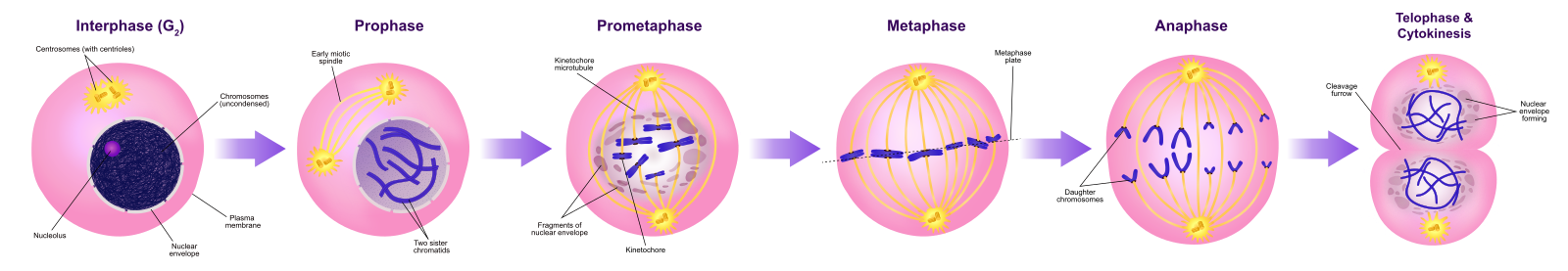
Prophase
Pendant la prophase, la « première phase », l’enveloppe nucléaire commence à se dissocier en petites vésicules, et les organelles membranaires (comme l’appareil de Golgi et le réticulum endoplasmique), se fragmentent et se dispersent vers les bords de la cellule. Le nucléole disparaît. Les centrosomes commencent à se déplacer vers les pôles opposés de la cellule. Les microtubules qui formeront le fuseau mitotique s’étendent entre les centrosomes, les éloignant les uns des autres à mesure que les fibres microtubulaires s’allongent. Les chromatides sœurs commencent à s’enrouler plus étroitement à l’aide des protéines condensines et deviennent visibles au microscope optique.
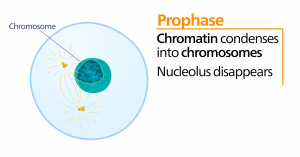
Prométaphase
Pendant la prométaphase, la « première phase de changement », de nombreux processus commencés en prophase continuent à progresser. Les restes de l’enveloppe nucléaire se fragmentent. Le fuseau mitotique continue à se développer alors que davantage de microtubules s’assemblent et s’étirent sur la longueur de l’ancienne zone nucléaire. Les chromosomes deviennent plus condensés et plus discrets. Chaque chromatide sœur développe une structure protéique appelée kinétochore dans la région centromérique.
Les protéines du kinétochore attirent et lient les microtubules du fuseau mitotique. Lorsque les microtubules du fuseau s’étendent à partir des centrosomes, certains de ces microtubules entrent en contact avec les cinétochores et s’y fixent fermement. Une fois qu’une fibre mitotique s’attache à un chromosome, ce dernier s’oriente jusqu’à ce que les cinétochores des chromatides sœurs fassent face aux pôles opposés. Finalement, toutes les chromatides sœurs seront attachées par leurs cinétochores aux microtubules des pôles opposés. Les microtubules du fuseau qui n’engagent pas les chromosomes sont appelés microtubules polaires. Ces microtubules se chevauchent à mi-chemin entre les deux pôles et contribuent à l’allongement de la cellule. Les microtubules astraux sont situés près des pôles, aident à l’orientation du fuseau et sont nécessaires à la régulation de la mitose.

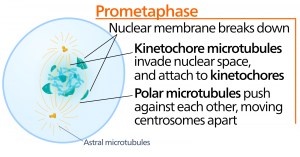
Métaphase
Pendant la métaphase, la « phase de changement », tous les chromosomes sont alignés dans un plan appelé plaque de métaphase, ou plan équatorial, à mi-chemin entre les deux pôles de la cellule. Les chromatides sœurs sont encore solidement attachées les unes aux autres par des protéines de cohésine. À ce moment, les chromosomes sont condensés au maximum.
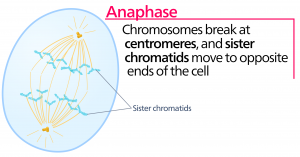
Anaphase
Pendant l’anaphase, la « phase ascendante », les protéines de cohésine se dégradent et les chromatides sœurs se séparent au centromère. Chaque chromatide, maintenant appelée chromosome, est tirée rapidement vers le centrosome auquel son microtubule est attaché. La cellule devient visiblement allongée (de forme ovale) alors que les microtubules polaires glissent les uns contre les autres au niveau de la plaque de métaphase où ils se chevauchent.
Telophase
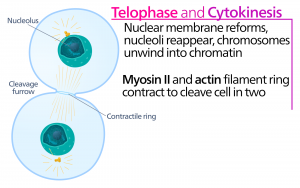
Pendant la télophase, la « phase de distance », les chromosomes atteignent les pôles opposés et commencent à se décondenser (se défaire), se relaxant dans une configuration chromatinienne. Les fuseaux mitotiques sont dépolymérisés en monomères de tubuline qui seront utilisés pour assembler les composants du cytosquelette de chaque cellule fille. Les enveloppes nucléaires se forment autour des chromosomes, et les nucléosomes apparaissent dans la zone nucléaire.
Cytokinèse
La cytokinèse, ou « mouvement cellulaire », est la deuxième étape principale de la phase mitotique au cours de laquelle la division cellulaire est achevée via la séparation physique des composants cytoplasmiques en deux cellules filles. La division n’est pas terminée tant que les composants cellulaires n’ont pas été divisés et complètement séparés en deux cellules filles. Bien que les étapes de la mitose soient similaires pour la plupart des eucaryotes, le processus de cytokinèse est assez différent pour les eucaryotes qui ont des parois cellulaires, comme les cellules végétales.
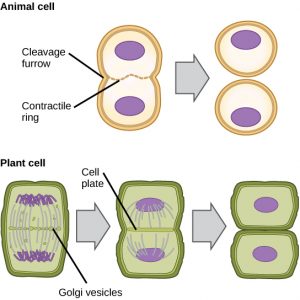
Dans les cellules comme les cellules animales qui n’ont pas de parois cellulaires, la cytokinèse suit le début de l’anaphase. Un anneau contractile composé de filaments d’actine se forme juste à l’intérieur de la membrane plasmique au niveau de l’ancienne plaque de métaphase (figure 12). Les filaments d’actine tirent l’équateur de la cellule vers l’intérieur, formant une fissure. Cette fissure, ou « fente », s’appelle le sillon de clivage. Le sillon se creuse au fur et à mesure que l’anneau d’actine se contracte, et finalement la membrane est clivée en deux.
Dans les cellules végétales, une nouvelle paroi cellulaire doit se former entre les cellules filles. Pendant l’interphase, l’appareil de Golgi accumule des enzymes, des protéines structurelles et des molécules de glucose avant de se briser en vésicules et de se disperser dans toute la cellule en division (figure 12). Pendant la télophase, ces vésicules de Golgi sont transportées sur des microtubules pour former un phragmoplaste (une structure vésiculaire) au niveau de la plaque métaphasique. Là, les vésicules fusionnent et coalescent du centre vers les parois cellulaires ; cette structure est appelée plaque cellulaire. Au fur et à mesure que d’autres vésicules fusionnent, la plaque cellulaire s’agrandit jusqu’à ce qu’elle fusionne avec les parois cellulaires à la périphérie de la cellule. Les enzymes utilisent le glucose qui s’est accumulé entre les couches membranaires pour construire une nouvelle paroi cellulaire. Les membranes de Golgi deviennent des parties de la membrane plasmique de chaque côté de la nouvelle paroi cellulaire.
Résumé de la mitose et de la cytokinèse
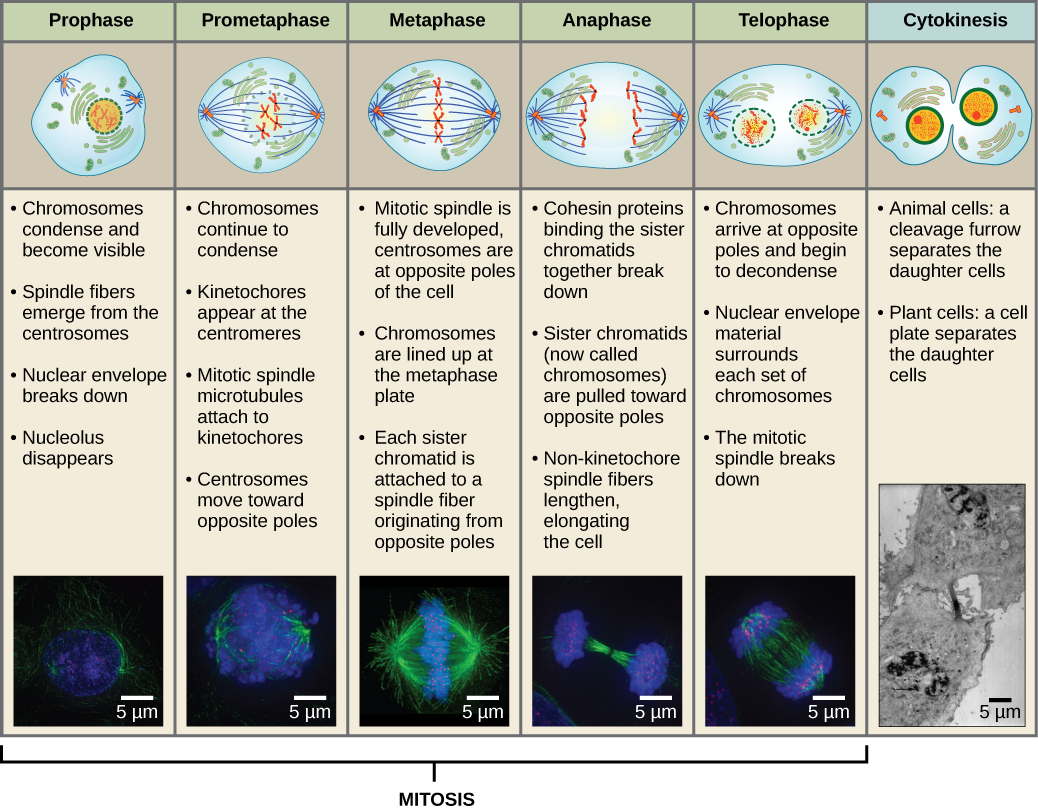
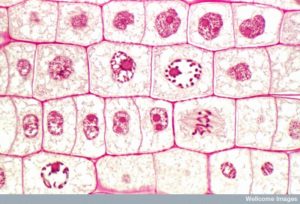
Figure 13 La mitose est divisée en cinq étapes-prophase, prométaphase, métaphase, anaphase et télophase. Les photos du bas ont été prises par microscopie à fluorescence de cellules artificiellement colorées par des colorants fluorescents : la fluorescence bleue indique l’ADN (chromosomes) et la fluorescence verte indique les microtubules (appareil fusiforme). (crédit « dessins de mitose » : modification du travail de Mariana Ruiz Villareal ; crédit « micrographies » : modification du travail de Roy van Heesbeen ; crédit « micrographie de cytokinèse » : Centre Wadsworth/Département de la santé de l’État de New York ; données de barre d’échelle de Matt Russell)
Phase G0
Pas toutes les cellules adhèrent au schéma classique du cycle cellulaire dans lequel une cellule fille nouvellement formée entre immédiatement en interphase, suivie de près par la phase mitotique. Les cellules en phase G0 ne se préparent pas activement à se diviser. La cellule est dans un stade quiescent (inactif), ayant quitté le cycle cellulaire. Certaines cellules entrent temporairement en G0 jusqu’à ce qu’un signal externe déclenche le début de la G1. D’autres cellules qui ne se divisent jamais ou rarement, comme les cellules musculaires cardiaques et nerveuses matures, restent en G0 de façon permanente).