A sejtosztódásnak két fő típusa van az eukariótáknál: a mitózis és a meiózis. A mitózis a növekedéshez és gyógyuláshoz szükséges új testsejtek előállítására szolgál, míg a meiózist a nemi sejtek (petesejtek és spermiumok) előállítására használják. A meiózist egy későbbi fejezetben tárgyaljuk.
A sejtciklus a sejtnövekedést és a sejtosztódást magában foglaló események rendezett sorozata, amely a mitózis révén két új leánysejtet hoz létre. A sejtciklus hossza még az egyes szervezetek sejtjein belül is nagyon változó. Az emberben a sejtek cserélődésének gyakorisága a korai embrionális fejlődés során néhány órától a hámsejtek esetében átlagosan két-öt napig, vagy az egész emberi életen át osztódás nélkül eltöltött időig terjed az olyan specializált sejteknél, mint a kérgi idegsejtek vagy a szívizomsejtek. Az is változó, hogy egy sejt mennyi időt tölt a sejtciklus egyes fázisaiban. Ha gyorsan osztódó emlőssejteket tenyésztenek kultúrában (a testen kívül, optimális növekedési körülmények között), a ciklus hossza körülbelül 24 óra. A sejtciklus eseményeinek időzítését a sejten belüli és kívüli mechanizmusok egyaránt szabályozzák.
A sejtek a sejtosztódás útján a növekedés, a DNS-replikáció és az osztódás pontosan időzített és gondosan szabályozott szakaszainak sorozatán haladnak keresztül, amelyekből két genetikailag azonos sejt jön létre. A sejtciklusnak két fő fázisa van: az interfázis és a mitotikus fázis (1. ábra). Az interfázis során a sejt növekszik és a DNS replikálódik. A mitotikus fázis során a replikált DNS és a citoplazma tartalma elválik egymástól, és a sejt osztódik.
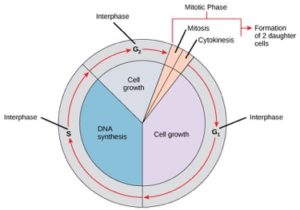
Az interfázis során a sejt normális folyamatokon megy keresztül, miközben a sejtosztódásra is felkészül. Ahhoz, hogy a sejt az interfázisból a mitotikus fázisba lépjen, számos belső és külső feltételnek kell teljesülnie. Az interfázis három szakaszát G1-nek, S-nek és G2-nek nevezzük.
G1 fázis (első rés)
Az interfázis első szakaszát G1 fázisnak (első rés) nevezzük, mert mikroszkopikus szempontból kevés változás látható. A G1 szakasz alatt azonban a sejt biokémiai szinten meglehetősen aktív. A sejt felhalmozza a kromoszómális DNS és a kapcsolódó fehérjék építőköveit, valamint elegendő energiatartalékot halmoz fel ahhoz, hogy a sejtmagban az egyes kromoszómák replikációjának feladatát elvégezze.
S fázis (DNS-szintézis)
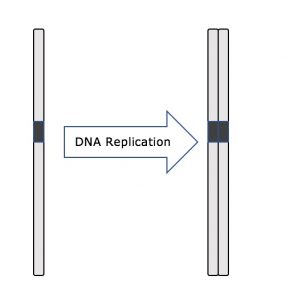
Az interfázis során a sejtmag DNS-e félig kondenzált kromatin konfigurációban marad. Az S fázisban a DNS-replikáció olyan mechanizmusok révén folytatódhat, amelyek eredményeként azonos DNS-molekulapárok – testvérkromatidák – képződnek, amelyek szilárdan kapcsolódnak a centromerikus régióhoz (2. ábra).
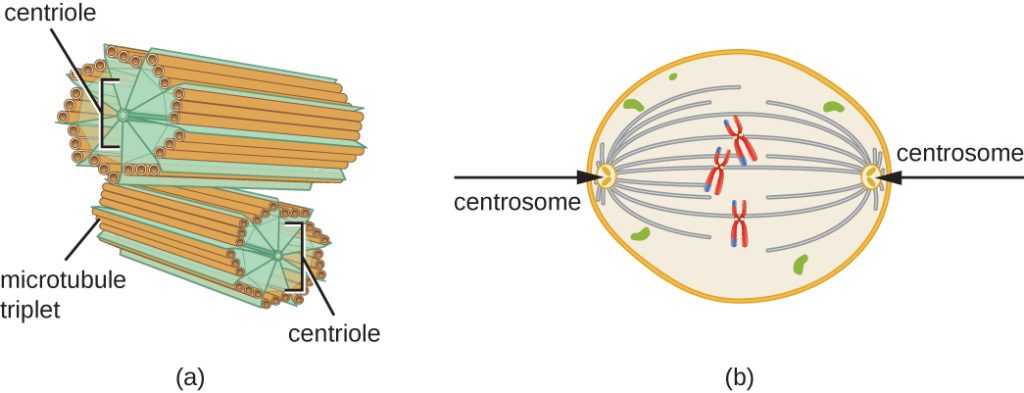
A centroszóma is megkettőződik az S fázis során. A két centroszóma adja majd a mitotikus orsót, azt az apparátust, amely a kromoszómák mozgását hangszereli a mitózis során. Az állati sejtek centroszómáihoz az egyes állati sejtek középpontjában egy pár pálcaszerű objektum, a centriolák kapcsolódnak, amelyek derékszögben állnak egymással. A centriolák segítenek a sejtosztódás megszervezésében. A centriolák más eukarióta fajok, például a növények és a legtöbb gomba centroszómáiban nincsenek jelen.
G2 fázis (második rés)
A G2 fázisban a sejt feltölti energiaraktárait és szintetizálja a kromoszómamanipulációhoz szükséges fehérjéket. Néhány sejtorganellum megkettőződik, és a citoszkeleton lebomlik, hogy a mitotikus fázishoz szükséges erőforrásokat biztosítsa. A G2 során további sejtnövekedés következhet be. A mitotikus fázis végső előkészületeit be kell fejezni, mielőtt a sejt beléphet a mitózis első szakaszába.
A mitotikus fázis
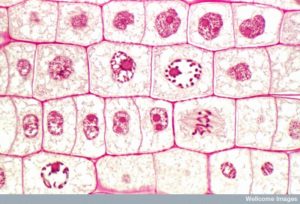
A két leánysejt létrehozásához a sejtmag és a citoplazma tartalmát kell megosztani. A mitotikus fázis egy többlépcsős folyamat, amelynek során a megkettőzött kromoszómák összehangolódnak, szétválnak és a sejt ellentétes pólusaira kerülnek, majd a sejt két új, azonos leánysejtre oszlik. A mitotikus fázis első része, a mitózis öt szakaszból áll, amelyek a sejtmag osztódását valósítják meg (5. ábra). A mitotikus fázis második része, az úgynevezett citokinézis, a citoplazmatikus komponensek fizikai szétválasztása két leánysejtre. Bár a mitózis szakaszai a legtöbb eukarióta esetében hasonlóak, a sejtfallal rendelkező eukarióták, például a növényi sejtek esetében a citokinézis folyamata teljesen eltérő.
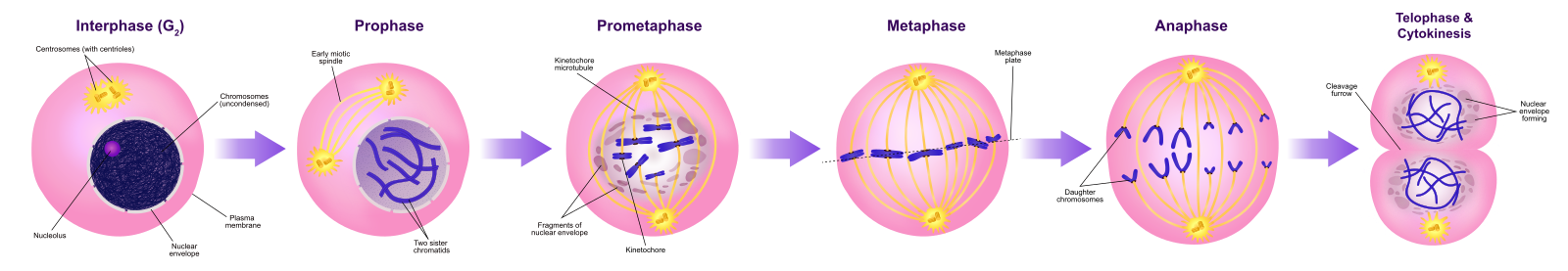
Profázis
A profázis, az “első fázis” során a magburkolat elkezd disszociálni kis vezikulákra, és a membránorganellák (például a Golgi-apparátus és az endoplazmatikus retikulum) feldarabolódnak és szétszóródnak a sejt szélei felé. A nukleolus eltűnik. A centroszómák elkezdenek a sejt ellentétes pólusai felé mozogni. A mitotikus orsót alkotó mikrotubulusok a centroszómák közé nyúlnak, távolabb tolva őket egymástól, ahogy a mikrotubulusrostok meghosszabbodnak. A testvérkromatidák a kondenzin fehérjék segítségével elkezdenek szorosabban tekeredni, és fénymikroszkóp alatt láthatóvá válnak.
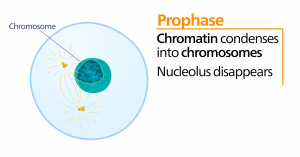
Prometafázis
A prometafázis, az “első változási fázis” során számos, a profázisban megkezdett folyamat folytatódik. A magburkolat maradványai feldarabolódnak. A mitotikus orsó tovább fejlődik, ahogy egyre több mikrotubulus gyűlik össze és feszül ki a korábbi magterület hosszában. A kromoszómák egyre sűrűbbé és diszkrétebbé válnak. Minden egyes testvérkromatidán a centromerikus régióban kinetochornak nevezett fehérjeszerkezet alakul ki.
A kinetokór fehérjéi vonzzák és megkötik a mitotikus orsó mikrotubulusait. Ahogy az orsó mikrotubulusai kinyúlnak a centroszómákból, e mikrotubulusok egy része érintkezik a kinetokórákkal, és szilárdan kötődik hozzájuk. Amint egy mitotikus szál egy kromoszómához kapcsolódik, a kromoszóma addig orientálódik, amíg a testvérkromatidák kinetokórái az ellentétes pólusok felé néznek. Végül az összes testvérkromatida a kinetochorjaikon keresztül az ellentétes pólusokról származó mikrotubulusokhoz fog kapcsolódni. Az orsó mikrotubulusait, amelyek nem kapcsolódnak a kromoszómákhoz, poláris mikrotubulusoknak nevezzük. Ezek a mikrotubulusok a két pólus között félúton átfedik egymást, és hozzájárulnak a sejt megnyúlásához. Az asztrális mikrotubulusok a pólusok közelében helyezkednek el, segítik az orsó orientációját, és szükségesek a mitózis szabályozásához.

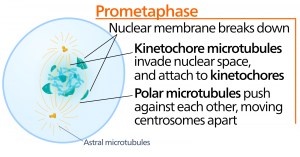
Metafázis
A metafázis, a “változási fázis” során az összes kromoszóma a sejt két pólusa között középen elhelyezkedő, metafázislemeznek vagy ekvatoriális síknak nevezett síkban igazodik. A testvérkromatidák még mindig szorosan kapcsolódnak egymáshoz a cohesin fehérjék segítségével. Ekkor a kromoszómák maximálisan kondenzáltak.
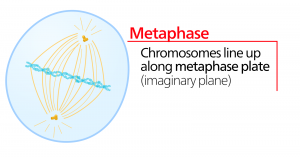
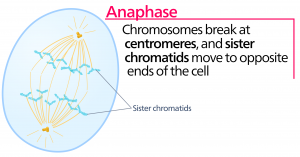
Anapfázis
Az anafázis, a “felfelé irányuló fázis” során a kohézinfehérjék lebomlanak, és a testvérkromatidák szétválnak a centromérnél. Mindegyik kromatidát, amelyet most már kromoszómának nevezünk, gyorsan a centroszóma felé húzza, amelyhez mikrotubulusa kapcsolódik. A sejt láthatóan megnyúlik (ovális alakúvá válik), ahogy a poláris mikrotubulusok egymáshoz csúsznak a metafázislemezen, ahol átfedik egymást.
Telophase
A telophase, a “távolsági fázis” során a kromoszómák elérik az ellentétes pólusokat, és elkezdenek dekondenzálódni (kibomlani), kromatinkonfigurációba lazulni. A mitotikus orsók depolimerizálódnak tubulin-monomerekké, amelyekből az egyes leánysejtek citoszkeletális komponensei összeállnak. A kromoszómák körül magburok alakul ki, és a nukleoszómák megjelennek a magterületen belül.
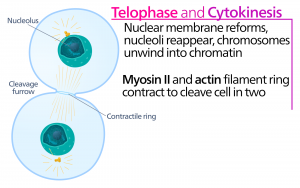
Citokinézis
A citokinézis, vagy “sejtmozgás” a mitotikus fázis második fő szakasza, amelynek során a sejtosztódás a citoplazmatikus összetevők fizikai szétválasztásával két leánysejtre fejeződik be. Az osztódás csak akkor fejeződik be, ha a sejtkomponensek szétválnak és teljesen szétválnak a két leánysejtre. Bár a mitózis szakaszai a legtöbb eukarióta esetében hasonlóak, a sejtfallal rendelkező eukarióták, például a növényi sejtek esetében a citokinézis folyamata teljesen eltérő.
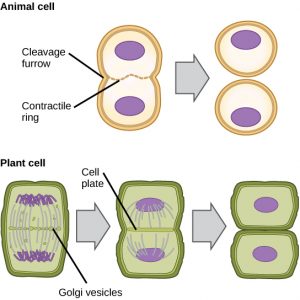
A sejtfalakkal nem rendelkező sejtekben, például az állati sejtekben a citokinézis az anafázis kezdetét követi. Egy aktin filamentumokból álló kontraktilis gyűrű alakul ki közvetlenül a plazmamembrán belsejében a korábbi metafázislemeznél (12. ábra). Az aktin filamentumok behúzzák a sejt egyenlítőjét befelé, hasadékot képezve. Ezt a hasadékot vagy “repedést” nevezzük hasadási barázdának. A barázda az aktingyűrű összehúzódásával egyre mélyül, és végül a membrán kettéhasad.
A növényi sejtekben a leánysejtek között új sejtfalnak kell kialakulnia. Az interfázis során a Golgi-apparátus enzimeket, szerkezeti fehérjéket és glükózmolekulákat halmoz fel, mielőtt vezikulákra bomlana és szétszóródna az osztódó sejtben (12. ábra). A telofázis során ezek a Golgi-vezikulák mikrotubulusokon szállítják őket, hogy a metafázislemezen fragmoplasztot (egy vezikuláris struktúrát) képezzenek. Ott a vezikulák összeolvadnak és összeolvadnak a középponttól a sejtfalak felé; ezt a struktúrát nevezzük sejtlemeznek. Ahogy egyre több vezikulum fuzionál, a sejtlemez megnagyobbodik, amíg a sejt perifériáján össze nem olvad a sejtfalakkal. A membránrétegek között felhalmozódott glükózt enzimek használják fel az új sejtfal felépítéséhez. A Golgi-membránok az új sejtfal mindkét oldalán a plazmamembrán részeivé válnak.
A mitózis és a citokinézis összefoglalása
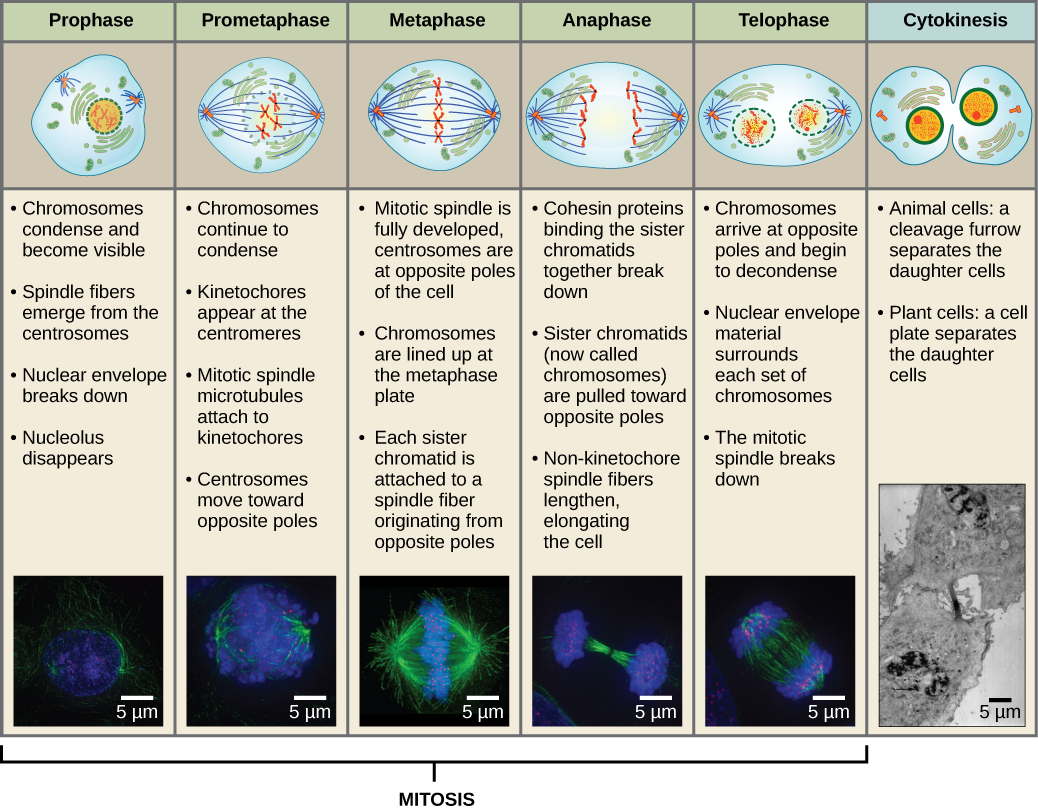
13. ábra A mitózis öt szakaszra oszlik: profázis, prometafázis, metafázis, anafázis és télofázis. Az alsó képeket fluoreszcens mikroszkópiával készítettük a fluoreszcens festékekkel mesterségesen megfestett sejtekről: a kék fluoreszcencia a DNS-t (kromoszómák), a zöld fluoreszcencia a mikrotubulusokat (orsóapparátus) jelzi. (credit “mitózis rajzok”: Mariana Ruiz Villareal munkájának módosítása; credit “mikrográfok”: Roy van Heesbeen munkájának módosítása; credit “citokinézis mikrográfok”: Wadsworth Center/New York Állami Egészségügyi Minisztérium; a méretarányos adatok Matt Russelltől származnak)
G0 fázis
Nem minden sejt követi a klasszikus sejtciklus-mintát, amelyben az újonnan keletkezett leánysejt azonnal belép az interfázisba, amelyet szorosan követ a mitotikus fázis. A G0 fázisban lévő sejtek nem készülnek aktívan osztódásra. A sejt nyugalmi (inaktív) szakaszban van, miután kilépett a sejtciklusból. Egyes sejtek átmenetileg belépnek a G0 fázisba, amíg egy külső jel el nem indítja a G1 fázis kezdetét. Más sejtek, amelyek soha vagy ritkán osztódnak, mint például az érett szívizom- és idegsejtek, tartósan a G0 fázisban maradnak).