Introduction
Charophytes or basal Streptophytes; (Becker and Marin, 2009; Leliaert et al., 2012) a ma élő édesvízi és szárazföldi zöldalgák változatos taxonómiai választékát alkotják, amelyek az egysejtű, fonalas és “parenchimatikus” formák széles skáláját mutatják (Graham, 1993; Lewis és McCourt, 2004). A charofiták egy ősi vonala 450-500 millió évvel ezelőtt jelent meg és telepedett meg a szárazföldön. Ezek az organizmusok alkalmazkodtak a szárazföldi körülményekhez, képessé váltak a túlélésre és a szaporodásra, amikor teljesen ki voltak téve a légkörnek, és egyes tagjai végül szárazföldi növényekké fejlődtek (Zhong et al., 2013; Delwiche és Cooper, 2015). A zöld növényeknek ez a “terrestrializációja” olyan kulcsfontosságú biológiai eseményt jelentett, amely örökre megváltoztatta a bolygó biogeokémiáját és természetrajzát. A szárazföldi növények ezt követő elterjedése megváltoztatta a légköri és a talajviszonyokat, és tovább módosította azokat, valamint lehetővé tette más változatos életformák megjelenését a szárazföldön. A szárazföldi növények evolúciója az emberi történelmet is átalakította, a legjelentősebb mértékben a mezőgazdaság újítása és a modern emberi civilizáció ebből következő megteremtése révén. Részben evolúciós jelentőségük miatt a karofiták az elmúlt évtizedekben jelentős figyelmet kaptak a növénybiológusoktól (Pickett-Heaps és Marchant, 1972; Pickett-Heaps, 1975; Mattox és Stewart, 1984; Becker és Marin, 2009; Harholt et al., 2016). Ahogy azonban további új adatok gyűltek össze ezen algák biológiájával kapcsolatban, fontos modellé váltak a biokémia, a sejtbiológia, a fejlődésbiológia, az ökológia és egyre inkább a molekuláris biológia alapvető jelenségeinek megértéséhez (Delwiche és Cooper, 2015; Lemieux et al., 2016). A “Charophytes” című Frontiers in Plant Science sorozat a növénybiológiai kutatások számos konkrét területén illusztrálja ezeknek az organizmusoknak a jelentőségét. Ez a mini-áttekintés kiemeli a karofiták mint modellorganizmusok tulajdonságait a kutatás különböző területein. Emellett remélhetőleg ösztönzést nyújt a tudósok új generációja számára, hogy kiterjesszék ezen algák használatát az alapkutatásban, és kezdeményezzék más taxonok szűrését, ami viszont új modellcharofiták azonosításához és felhasználásához vezethet.
A charofiták “sora”
Az 1970-es és 1980-as években az ultrastrukturális (pl., sejtosztódási mechanizmus, flagelláris apparátus felépítése) és biokémiai (pl. enzimprofilok) adatok voltak a fő kritériumai annak, hogy egy zöldalgát a charophyták vonalába soroljanak (Mattox és Stewart, 1984; Lewis és McCourt, 2004; Leliaert et al., 2012). Az 1990-es évektől napjainkig a kloroplasztisz- és nukleáris genomok és transzkriptomok molekuláris elemzéseire összpontosító vizsgálatok megerősítették és finomították a korábbi rendszertani és filogenetikai sémákat (Timme és mtsai., 2012; Delwiche és Cooper, 2015). Ezt kiegészítették továbbá a karofita sejtfalak és a hormonok bioszintetikus és jelátviteli útvonalainak részletes biokémiai, immunológiai és sejtbiológiai alapú elemzései (Popper és Fry, 2003; Popper, 2008; Sørensen et al., 2010, 2011, 2012; Zhang és van Duijn, 2014; Ju et al., 2015; O’Rourke et al., 2015). A létező charofiták más zöldalga-taxonokhoz képest viszonylag alacsony százalékos diverzitást mutatnak, és 13 családot és 122 nemzetséget foglalnak magukban (Becker és Marin, 2009; Leliaert et al., 2012). A jelenlegi filogenetikai vélemények a charophytákat hat osztályba sorolják (1. ábra; Delwiche és Cooper, 2015). A bazális osztályt, a Mesostigmatophyceae-t egyetlen ismert nemzetség, a Mesostigma képviseli. Ez az alga egy egysejtű biflagellata, amely egyedülállóan aszimmetrikus alakú (azaz lapított mentőcsónakhoz hasonló), és díszesen kialakított pikkelyek rétegei borítják (Manton és Ettl, 1965; Becker és mtsai., 1991; Domozych és mtsai., 1991, 1992). A második osztály, a Chlorokybophyceae, szintén egyetlen ismert típusból, a Chlorokybus atmophyticusból áll. Ez a ritka alga szarkinoid sejtcsomagot alkot, amelyet vastag, gélszerű borítás vesz körül (Rogers et al., 1980). Meg kell jegyezni, hogy az alternatív filogenezisek a Mesostigmát és a Chlorokybust testvérvonalakként helyezik el, amelyek együtt az összes többi Streptophytával testvérek (Rodríguez-Ezpeleta et al., 2007). A hat osztályos filogeniában a harmadik osztály, a Klebsormidiophyceae három nemzetségből áll, amelyek egyszerű, elágazás nélküli fonalakat alkotnak (Sluiman et al., 2008), amelyek gyakran a különböző szárazföldi élőhelyek felszínén növekvő “biológiai kérgek” tagjai. Az ebbe az osztályba tartozó számos faj még a sivatagi kéreg részeként is képes ellenállni a jelentős kiszáradási stressznek (Mikhailyuk et al., 2008, 2014; Holzinger és Karsten, 2013); mások az alacsony hőmérsékletű környezetekkel járó kiszáradási stresszel szemben is toleránsak (Stamenkovic et al., 2014; Herburger és Holzinger, 2015). Ez az első három osztály alkotja a “korai divergens” charophytákat. A “késői divergens” vonal szintén három osztályt foglal magában. A Charophyceae, vagyis a kőtörzsűek általában édesvízi ökoszisztémákban fordulnak elő, és makroszkopikus, többsejtű talirokkal rendelkeznek, amelyek kivételesen hosszúkás sejtekből álló, elágazó szálakból álló aggregátumokból állnak (Lewis és McCourt, 2004). E csoport tagjai oogámián alapuló szexuális szaporodással rendelkeznek, amely magában foglalja a mozgékony spermiumokat és a nem mozgékony petéket, amelyek mindkettő többsejtű gametangiumokban termelődik. A Coleochaetophyceae csoportba olyan taxonok tartoznak, amelyeknek többsejtű, elágazó fonalas vagy parenchimaszerű fonalból álló talaja van (Graham, 1993; Delwiche et al., 2002). Ezek az organizmusok is megkülönböztetett oogámia-alapú szexuális szaporodást mutatnak. A Coleochaetophyceae jellemzően az édesvízi és szárazföldi élőhelyek határán, gyakran epifitaként fordulnak elő. Végül a charophyták legnagyobb és legváltozatosabb csoportja, a Zygnematophyceae egysejtűekből és elágazás nélküli fonalakból áll (Gontcharov et al., 2003; Guiry, 2013). E csoport megkülönböztető jellemzője a konjugáción alapuló szexuális szaporodás jelenléte, amely komplex sejt-sejt jelátvitelt és adhéziót igényel (Abe et al., 2016). A zigmatofíliás algák gyakori lakói az édesvízi élőhelyeknek, néha látványos efemer virágzásokban, valamint nedves szárazföldi szubsztrátumokban is előfordulnak.

1. ábra. Kiterjedt Charophyták. Az extant Charophytákat korai és késői divergáló taxonokra osztjuk. A korai divergáló taxonok közé tartoznak a Mesostigmatophyceae, a Chlorokybophyceae és a Klebsormidiophyceae, a késői divergáló taxonok közé pedig a Charophyceae, a Coleochaetophyceae és a Zygnematophyceae. Az egyes csoportok képviselői a Mesostigma (Mesostigmatophyceae), Chlorokybus atmophyticus (Chlorokybophyceae), Klebsormidium flaccidum (Klebsormidiophyceae), Chara (Charophyceae), Coleochaete scutata (Coleochaetophyceae) és Micrasterias (Zygnematophyceae).
A charophyták mint modellszervezetek
Egyes charophytákat széles körben használtak modellszervezetként az alapvető biológiai folyamatok vizsgálatában. Kicsi és egyszerű talijuk (pl. a szárazföldi növényekhez képest) és könnyű kísérleti manipulálhatóságuk csak két olyan tulajdonságuk, amely vonzó modellorganizmusokká teszi őket. A legújabb bizonyítékok azt is kimutatták, hogy számos charofita számos figyelemre méltóan hasonló tulajdonsággal rendelkezik, mint a szárazföldi növények, beleértve számos növekedésszabályozó bioszintetikus útvonalának (2. ábra; Boot et al., 2012; Hori et al., 2014; Wang et al., 2014, 2015; Holzinger és Becker, 2015; Ju et al., 2015) és többféle sejtfalpolimer jelenlétét (Popper, 2008; Sørensen et al., 2010, 2011, 2012; Mikkelsen et al., 2014). Ez a két tulajdonság tette a karofitákat hatékonnyá olyan kutatási területeken, mint a növényi molekuláris fejlődés és a stresszfiziológia. Bár számos charofitát használtak fel a biológiai vizsgálatok széles skáláján, a következő taxonok a legjelentősebbek, mivel a tanulmányok több területén is széles körben használják őket modellként:
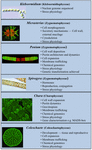
2. ÁBRA. A jelenlegi modell Charophyták. Számos Charophytát, köztük a Klebsormidiumot, a Micrasteriast, a Peniumot, a Spirogyrát, a Charát és a Coleochaete-t használják modellként; mindegyiknek vannak olyan sajátos tulajdonságai, amelyek különösen alkalmassá teszik bizonyos élettani és biokémiai folyamatok modelljévé.
Desmidák: A szimmetrikus modellek
A Zygnematophyceae-ről nemrégiben kimutatták, hogy valószínűleg a szárazföldi növények legközelebbi élő ősei (Wodniok és mtsai, 2011; Delwiche és Cooper, 2015). Az inkluzív placoderm desmid csoport egysejtű tagjai fontos modellekké váltak a növényi sejtbiológia és fejlődés számos alapvető elvének megvilágításában. A nagy sejtméret, a figyelemre méltó szimmetria/forma, a határozott sejtfal-architektúra és a kidolgozott endomembrán/citovázrendszer csak néhány azon jellemzők közül, amelyek kiváló sejtrendszerré teszik őket a sejtfiziológia és a fejlődés tanulmányozására. A közelmúltban létrehozott stabil transzformált vonalak és a kiválasztott desmidák hamarosan szekvenált genomjai/transzkriptomjai tovább növelték értéküket a botanikai vizsgálatokban. A desmidok közül két taxon, a Micrasterias és a Penium vált a legjobban tanulmányozott taxonná.
A Micrasterias az elmúlt 50 évben egyedülálló tulajdonságainak köszönhetően a legnépszerűbb desmid volt a sejtbiológiai kutatásokban (Meindl, 1993; Lutz-Meindl, 2016). A Micrasterias sejt kétoldali szimmetriát mutat, amelyet gyakran látványosan tagolt (azaz multilobális) perifériával emelnek ki. A sejtosztódás során keletkező új leányfélsejtek nem rendelkeznek ezzel az összetett morfológiával, hanem inkább gömb alakúak. Egy összetett poszt-citokinetikus fejlődési program, amely több, jól összehangolt szubcelluláris mechanizmust alkalmaz, később a leánysejtek terjeszkedése során létrehozza a multilobális fenotípust. Ez a program felelős a polimerek lerakódásáért is az elsődleges és a másodlagos sejtfal létrehozásához. Ezen események középpontjában a Golgi-testek és a kapcsolódó vezikulák nagy hálózata áll, amelyek egy összetett szekréciós mechanizmust eredményeznek. Ez a folyamat viszont specifikus sejtfelszíni helyekre irányul, amelyek többpólusú sejttágulást és a sejtfal makromolekulák és extracelluláris nyálkaanyag egyidejű szekrécióját hozzák létre (Kim és mtsai., 1996; Lutz-Meindl és Brosch-Salomon, 2000; Oertel és mtsai., 2004; Aichinger és Lutz-Meindl, 2005). A szekréciós komponensek pontos sejtfelszíni lokalizációkra történő eljuttatásához egy bonyolult aktinomiozin-alapú citomotil rendszerre van szükség, amelyet számos jelátviteli kaszkád vezérel (Meindl és mtsai., 1994; Oertel és mtsai., 2003). A sejtmorfogenezis továbbá rendkívül érzékeny a külső stresszre (pl. oxidatív stressz, sótartalom, nehézfémek), ami a sejtdifferenciálódás jelentős változásaihoz vezet (Darehshouri et al., 2008; Affenzeller et al., 2009; Andosch et al., 2012). A Micrasterias kiváló organizmus ezen szubcelluláris jellemzők elemzésére, mivel könnyen karbantartható és manipulálható a laboratóriumban, és különösen alkalmas a nagy felbontású képalkotás megszerzésére fény- és elektronmikroszkópiával, beleértve az immuncitokémiát, a fókuszált ionnyaláb-pásztázó elektronmikroszkópiát (FIB-SEM) és az elektron energiaveszteség spektroszkópiás (EELS) képalkotást (Lutz-Meindl, 2007; Eder és Lutz-Meindl, 2008, 2010; Wanner et al., 2013; Lutz-Meindl et al., 2015). Továbbá a kezdeti molekuláris elemzések, beleértve a transzformált sejtvonalak előállítását is, továbbfejlesztették a Micrasterias felhasználását, különösen a sejtfal feldolgozásának molekuláris dinamikájában (Vannerum et al., 2010, 2011, 2012).
Az elmúlt évtizedben a desmid, Penium margaritaceum, szintén értékes modellorganizmussá vált (Domozych et al., 2009, 2014; Sørensen et al., 2014; Rydahl et al., 2015; Worden et al., 2015). A Micrasteriasszal ellentétben a Penium egyszerű hengeres alakú, csak elsődleges sejtfallal rendelkezik, és a sejtfelszín két specifikus lokalizációján lerakódnak a falpolimerek a terjeszkedés során (Domozych és mtsai., 2011). Ez a viszonylagos egyszerűség rendkívül vonzó a növényi sejtfejlődés alapvető elveinek feltárásához, beleértve a sejtfal kialakulását, a sejttágulást és a szekréció dinamikáját (Domozych et al., 2005, 2014; Ochs et al., 2014). Egyik fő tulajdonsága, hogy a sejtfal polimerjei hasonlóak a számos szárazföldi növényben jelenlévőkhöz (pl. cellulóz, pektinek, hemicellulózok), és hogy ezek a polimerek monoklonális antitestekkel jelölhetők. Az élő sejtek jelölése után ezek visszahelyezhetők a kultúrába, ahol a későbbi sejttágulás és fallerakódási események nyomon követhetők (Domozych és mtsai., 2009; Rydahl és mtsai., 2015). A penium a laboratóriumban is könnyen fenntartható, és gyors növekedési sebessége pontosan ellenőrzött körülmények között kiváló mintává teszi számos kémiai hatóanyag nagyméretű, egyidejű mikroszűrésére (multi-well lemezekben történő növesztéssel) és a terjeszkedésre/falfejlődésre gyakorolt specifikus hatásuk értékelésére (Worden et al., 2015). Ez jelentősen segíti a specifikus szubcelluláris komponensek és folyamatok szerepének feltárását a terjeszkedésben/differenciálódásban. A közelmúltban a stabilan transzformált vonalak izolálása (Sørensen et al., 2014) tovább növelte ennek az algának a potenciálját a jövőbeli növényi sejtes vizsgálatokban.
Chara és Nitella: Cellular Giants for Plant Physiology
A Chara és Nitella (Charophyceae) fajokat régóta használják mintaként számos biológiai vizsgálathoz, különösen azokhoz, amelyek a sejtdinamikával, a terjeszkedéssel és a citoplazma áramlásával foglalkoznak (Green, 1954; Probine és Preston, 1962). Makroszkopikus tallijukat a csomópontok, ahol az elágazások és a gametangiumok keletkeznek, és a kivételesen megnyúlt sejtekből álló belső csomószakaszok különböztetik meg. Az internodális sejtek a szubcelluláris komponensek egyértelműen észrevehető rétegződését mutatják, ahol a helikális irányultságú kloroplasztiszok egy álló kéregállományt határoznak meg, amely egy endoplazmát vesz körül, ahol a leggyorsabban rögzített aktinomjozin által generált citoplazmaáramlás történik. A citoplazma változatos endomembrán komponenseket tartalmaz, amelyek dinamikus membránforgalom-hálózatokban vesznek részt (pl. endocitózis és exocitózis), amelyeket komplex jelátviteli kaszkádok irányítanak (Foissner és Wastenys, 2000, 2012, 2014; Sommer és mtsai., 2015; Foissner és mtsai., 2016). Az internodális sejtek cellulózban és pektinben, homogalakturonánban (HG) gazdag sejtfalakkal rendelkeznek. A turgor a terjeszkedés fő hajtóereje, a pektin pedig valószínűleg a fal terjeszkedését szabályozó teherviselő komponens. A HG ciklikus, nem enzimatikus modulációja és egyidejű kalcium-komplexképzése az a mechanizmus, amely a sejttáguláshoz vezető falmoduláció szabályozásáért felelős (Proseus és Boyer, 2007, 2008, 2012; Boyer, 2016). A Chara és a Nitella egyik technikai előnye a sejtkutatásban, hogy az internodális sejtek egyenként eltávolíthatók; az endoplazma és a sejtfalak “szétválaszthatók”, majd akelluláris rendszerként használhatók kísérleti manipulációhoz.
A többi taluszkomponens, például a rizoidok a citoplazma szervezett poláris szerveződésével rendelkeznek. Ezek a sejtek a csúcson elhelyezkedő, üledékes, ásványi anyagokban gazdag statolitokat tartalmaznak, amelyek a gravitáció érzékelésében működnek (Hodick et al., 1998; Braun, 2002; Braun és Limbach, 2006). Ez a tulajdonság tette a Chara-t az első charofitává az űrben. A TEXUS rakéta parabolikus repülései során kimutatták, hogy a statolitok feszítőerőt gyakorolnak az aktin filamentumokra. Az erők egyensúlya (azaz a gravitáció és az aktin filamentumok ellenhatása) felelős a statolitok megfelelő pozíciójáért a rizoidban, amelyek viszont garantálják a gravitációs vektorra való reagálás képességét (Braun és Limbach, 2006). Lézercsipeszek és lassan forgó centrifugamikroszkópia segítségével kimutatták, hogy a statolitok leülepedése nem elegendő a rizoid csúcsának gravitációs elhajlásához. Inkább az ásványi anyagokban gazdag sztatolitoknak kell a plazmamembrán meghatározott régióira telepedniük ahhoz, hogy gravitróp morfológiai hatások érvényesüljenek (Braun, 2002). A Chara-t más jelenségek, többek között az elektrofiziológia és a hormonok szerepének és mozgásának tanulmányozására is használták (Belby, 2016).
Coleochaete for Developmental Studies and Pattern Development
A növény többsejtű talusának (azaz szövetének, szervének) kialakulásához pontos, térben szabályozott sejtosztódási síkokra van szükség, amelyeket a fejlődés során meghatározott pontokon kifejeződő gének többféle készlete szabályoz (Besson és Dumais, 2011; Umen, 2014). A 3 dimenziós térben és időben egyaránt irányított thallus morfogenezis a külső stresszfaktorok hatására is modulálódik. A növényekben ezeket a fejlődési programokat még összetettebbé teszi a sejtfalak jelenléte, amelyek nem teszik lehetővé a sejtvándorlást vagy a szövetek rugalmasságát. A talus alakjának és méretének megnyilvánulásában központi szerepet játszó specifikus események tisztázása gyakran nehéz a szárazföldi növényekben, ahol a specifikus sejtviselkedések felbontása gyakran rosszul megoldott, amikor rendkívül összetett és kiterjedt szövetekbe/szervekbe ágyazódnak. Az alapvető fejlődési jelenségek megértéséhez előnyös volt az egyszerűbb talus kialakítású organizmusok használata, és a charofita Coleochaete nemzetség egy ilyen példa. Egyes fajok (pl. C. orbicularis, C. scutatum) parenchymatikus diszkoid thalluszt hoznak létre, amely anti- vagy periklinális sejtosztódások és a legkülső sejtek későbbi terjeszkedésének kombinációjával növekszik kifelé (Brown et al., 1994; Cook, 2004). Bármely sejt osztódási síkja egyszerű szabályokat követ, amelyek a sejtméreten, a sejt alakján és a thalluszon belüli földrajzi elhelyezkedésen alapulnak (Dupuy és mtsai., 2010; Besson és Dumais, 2011). Ez a tulajdonság lehetővé teszi olyan matematikai modellek felépítését, amelyek segítségével értelmezhető a többsejtű morfogenezis (Domozych és Domozych, 2014; Umen, 2014), és amelyek kritikus betekintést nyújtanak a szervek és egész szervezetek evolúciójába, biomechanikájába és fiziológiájába. Továbbá néhány Coleochaete-fajjal kiterjedten tanulmányozták azokat a szerkezeti és funkcionális modulációkat, amelyek a talluszban aszályos körülmények között következnek be (Graham és mtsai., 2012), azaz a korai szárazföldi növények evolúciójának és a szárazság alatti növényi növekedés dinamikájának megértéséhez kulcsfontosságú jellemzőket.
A genomikai és génvizsgálatokból származó felismerések
A molekuláris eszközök sok más technológiához hasonlóan képesek olyan új információkat közvetíteni, amelyek számos kutatási terület, köztük az élettan és a fejlődés szempontjából fontosak. Például a MADS-box géncsalád tagjait izolálták és jellemezték három charophytából, a Chara globularisból, a Coleochaete scutatából és a Closterium peracerosum-strigosum-littorale-ból (Tanabe et al., 2005). A MADS-box gének expressziós mintázata a charophytákban arra utal, hogy a haploid fejlődésben és szaporodásban játszanak szerepet (Tanabe et al., 2005). Úgy gondolják, hogy a szárazföldi növények evolúciója során a diploid generációba rekrutálódtak, és a fejlődésben betöltött szerepük tovább diverzifikálódott (Tanabe et al., 2005). A mai napig a Klebsormidium flaccidum az egyetlen olyan karofita, amelyről közel teljes nukleáris genom áll rendelkezésre (Hori et al., 2014); bár a karofiták mind a hat osztályának képviselőiről szekvenálták a kloroplasztisz és a mitokondriális genomot, és néhány karofitáról EST-k (expresszált szekvenciacímkék) állnak rendelkezésre (Delwiche, 2016). A Klebsormidium genomjában már kimutatták számos növényi hormon és jelátviteli köztitermék szintéziséhez szükséges gének jelenlétét, a nagy fényintenzitással szembeni védekezési mechanizmusokat (Hori et al., 2014), valamint a IIb csoportú WRKY transzkripciós faktorok jelenlétét, amelyekről korábban úgy gondolták, hogy először a mohákban jelentek meg (Rinerson et al., 2015). Jelenleg több charofita genomszekvenálási projekt is folyamatban van (Delwiche, 2016), amelyek részletesebb összehasonlító elemzéseket tesznek lehetővé, nagyobb számú charofita bevonásával, és potenciálisan új betekintést adhatnak a fajok és gének közötti evolúciós kapcsolatokba. A karofiták egyedülálló helyzete a streptofiták legkorábban divergáló csoportjaként különösen hatékony eszközzé teszi őket az evolúciós-fejlődéstani vizsgálatokhoz és az olyan új technológiákhoz, mint például a stabil transzformáció közelmúltbeli elérése a Peniumban (Sørensen et al, (2014) lehetővé teszi egyes gének funkciójának feltárását.
A karofiták jövőbeni felhasználása modellszervezetként
A karofiták kutatásának kritikus következő lépése az inkluzív taxonok genomjának elemzése és a szárazföldi növények genomjával való összehasonlító vizsgálatok. Ez a munka nemcsak kulcsfontosságú betekintést nyújt majd azokba az adaptív mechanizmusokba, amelyeket a charofiták a szárazföldi élőhelyekre való megjelenésük és sikeres kolonizációjuk során fejlesztettek ki, hanem tovább finomítja és bővíti a jelenlegi modellorganizmusok felhasználását az alapvető növénykutatásban. Bár a molekuláris vizsgálatok számos kutatási területet támogatnak majd, két olyan terület, amely azonnal hasznot fog hozni, a sejtfal biológiája és a növekedésszabályozók dinamikája a növekedés és a fejlődés során. Az előbbi területen a karofiták számos alapvető, de még kevéssé megoldott jelenséget tisztázhatnak, beleértve a pektin- és sejtfalfehérje-bioszintézist, a falpolimerek szabályozott szekrécióját, lerakódását és a szekréciót követő modulációkat, valamint a növényi sejtek specifikus, interaktív membránforgalom-hálózatát. Ez utóbbiban a karofita modellek segítenének megfejteni az olyan hormonokhoz, mint az etilén és az auxin, kapcsolódó molekuláris jelátvitelt, sejtbiológiát és a növekedés és fejlődés dinamikáját. A charofitákkal végzett közelmúltbeli ökofiziológiai vizsgálatok szintén kritikus adatok potenciális bónuszát kínálják annak meghatározásához, hogy a növényi sejtek hogyan alkalmazkodnak a stresszhez, beleértve a kiszáradást és a sótűrést (Holzinger és Karsten, 2013; Pichrtová és mtsai., 2014; Herburger és mtsai., 2015; Holzinger és Becker, 2015; Holzinger és Pichrtová, 2016; Kondo és mtsai., 2016). A charofita-kutatás jövője valóban nagyon fényes, és következésképpen az egész növénybiológia számára áldásos lesz.”
A szerzők hozzájárulása
A felsorolt szerzők mindegyike jelentősen, közvetlenül és intellektuálisan hozzájárult a munkához, és jóváhagyta azt a publikáláshoz.
Conflict of Interest Statement
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségként értelmezhetők.
Acknowledgement
Ez a munka az NSF (USA) MCB collaborative grants 1517345 és 1517546 támogatásával készült.
Belby, M. J. (2016). Több léptékű characea kísérleti rendszer: a membrántranszporterek elektrofiziológiájától a sejtek közötti összeköttetésig, a citoplazmatikus áramlásig és az auxin metabolizmusig. Front. Plant Sci. 7:1052. doi: 10.3389/fpls.2016.01052
CrossRef Full Text | Google Scholar
Braun, M. (2002). A gravitáció észlelése specifikus plazmamembránterületeken megtelepedett statolitokat igényel a characea rhizoidokban és protonémákban. Protoplasma 219, 150-159. doi: 10.1007/s007090200016
CrossRef Full Text | Google Scholar
Braun, M., and Limbach, C. (2006). A characea algák rizoidjai és protonémái: modellsejtek a polarizált növekedés és a növényi gravitációérzékelés kutatásához. Protoplasma 229, 133-142. doi: 10.1007/s00709-006-0208-9
CrossRef Full Text | Google Scholar
Graham, L. E. (1993). A szárazföldi növények eredete. New York, NY: John Wiley & Sons, 287.
Google Scholar
Herburger, K., and Holzinger, A. (2015). A kallóz lokalizációja és mennyiségi meghatározása a Zygnema és Klebsormidium streptofita zöldalgákban: korreláció a kiszáradás-tűréssel. Plant Cell Physiol. 56, 2259-2270. doi: 10.1093/pcp/pcv139
CrossRef Full Text | Google Scholar
Lemieux, C., Otis, C., and Turmel, M. (2016). Streptofita zöldalgák összehasonlító kloroplasztiszgenom-elemzései jelentős szerkezeti változásokat tárnak fel a Klebsormidiophyceae, Coleochaetophyceae és Zygnematophyceae családokban. Front. Plant Sci. 7:697. doi: 10.3389/fpls.2016.00697
CrossRef Full Text | Google Scholar
Lutz-Meindl, U. (2007). Energiaszűrő transzmissziós elektronmikroszkópia alkalmazása képalkotásra és elemelemzésre növényi szervezetekben. Micron 38, 181-196. doi: 10.1016/j.micron.2006.03.017
CrossRef Full Text | Google Scholar
Meindl, U. (1993). Micrasterias sejtek mint a morfogenezis kutatásának modellrendszere. Microbiol. Rev. 57, 415-433.
Google Scholar
Pichrtová, M., Kulichova, J., and Holzinger, A. (2014). A nitrogénlimitáció és a lassú száradás sarkvidéki élőhelyekről származó konjugáló zöldalgák (Zygnematophyceae, Streptophyta) kiszáradás-toleranciáját indukálja. PLoS ONE 9:e113137. doi: 10.1371/journal.pone.011313137
CrossRef Full Text | Google Scholar
Pickett-Heaps, J. D., and Marchant, H. J. (1972). A zöld algák filogeniája: egy új javaslat. Cytobios 6, 255-264.
Google Scholar
Stamenkovic, M., Woelken, E., and Hanelt, D. (2014). A különböző földrajzi helyekről gyűjtött Cosmarium-törzsek (Zygnematophyceae, Streptiophyta) ultrastruktúrája fajspecifikus különbségeket mutat mind optimális, mind stresszhőmérsékleten. Protoplasma 251, 1491-1509.
Google Scholar