Eukaryoten kennen twee belangrijke typen celdeling: mitose en meiose. Mitose wordt gebruikt om nieuwe lichaamscellen te produceren voor groei en genezing, terwijl meiose wordt gebruikt om geslachtscellen (eicellen en zaadcellen) te produceren. Meiose wordt in een later hoofdstuk besproken.
De celcyclus is een geordende reeks gebeurtenissen waarbij sprake is van celgroei en celdeling die via mitose twee nieuwe dochtercellen oplevert. De lengte van de celcyclus is zeer variabel, zelfs binnen de cellen van een individueel organisme. Bij de mens varieert de frequentie van de celwisseling van enkele uren in de vroege embryonale ontwikkeling tot gemiddeld twee tot vijf dagen voor epitheelcellen, of tot een heel mensenleven zonder deling in gespecialiseerde cellen zoals corticale neuronen of hartspiercellen. Er is ook variatie in de tijd die een cel in elke fase van de celcyclus doorbrengt. Wanneer snel delende zoogdiercellen in cultuur worden gebracht (buiten het lichaam onder optimale groeiomstandigheden), bedraagt de lengte van de cyclus ongeveer 24 uur. De timing van de gebeurtenissen in de celcyclus wordt gecontroleerd door mechanismen die zowel intern als extern aan de cel zijn.
Cellen op weg naar celdeling doorlopen een reeks nauwkeurig getimede en zorgvuldig gereguleerde stadia van groei, DNA-replicatie en deling die twee genetisch identieke cellen opleveren. De celcyclus kent twee belangrijke fasen: de interfase en de mitotische fase (figuur 1). Tijdens de interfase groeit de cel en wordt het DNA gerepliceerd. Tijdens de mitotische fase worden het gerepliceerde DNA en de cytoplasmatische inhoud van elkaar gescheiden en deelt de cel zich.
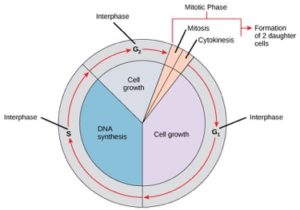
Tijdens de interfase ondergaat de cel normale processen terwijl hij zich ook voorbereidt op de celdeling. Om een cel van de interfase naar de mitotische fase te laten overgaan, moet aan veel interne en externe voorwaarden worden voldaan. De drie stadia van de interfase worden G1, S en G2 genoemd.
G1-fase (eerste kloof)
Het eerste stadium van de interfase wordt de G1-fase (eerste kloof) genoemd, omdat er microscopisch gezien weinig verandering zichtbaar is. Tijdens de G1-fase is de cel echter op biochemisch niveau behoorlijk actief. De cel accumuleert de bouwstenen van het chromosomale DNA en de bijbehorende eiwitten en bouwt voldoende energiereserves op om de taak van het repliceren van elk chromosoom in de celkern te voltooien.
S-fase (Synthese van DNA)
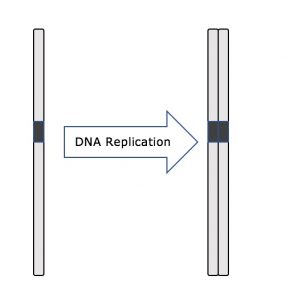
Tijdens de interfase blijft het kern-DNA in een semi-gecondenseerde chromatineconfiguratie. In de S-fase kan DNA-replicatie plaatsvinden via mechanismen die resulteren in de vorming van identieke paren DNA-moleculen – zustercromatiden – die stevig aan de centromeerregio zijn gehecht (figuur 2).
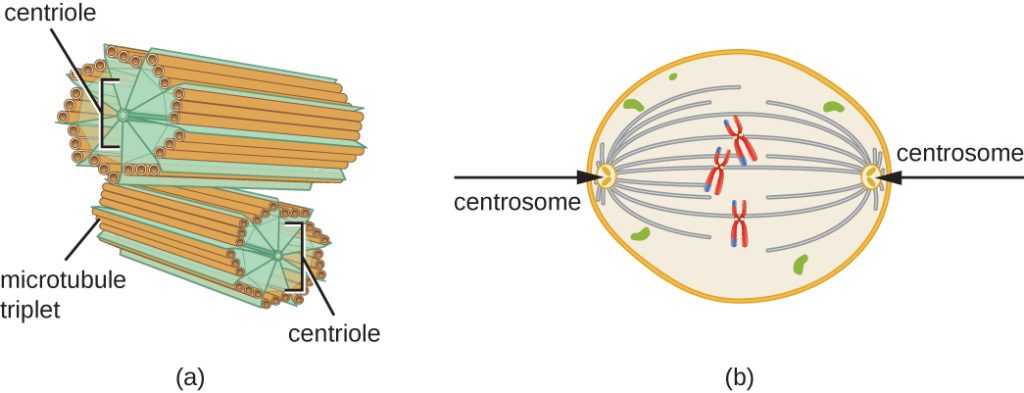
Het centrosoom wordt ook gedupliceerd tijdens de S-fase. Uit de twee centrosomen ontstaat de mitotische spindel, het apparaat dat tijdens de mitose de beweging van de chromosomen orkestreert. In het centrum van elke dierlijke cel zijn de centrosomen van dierlijke cellen geassocieerd met een paar staafachtige objecten, de centriolen, die loodrecht op elkaar staan. Centriolen helpen de celdeling te organiseren. Centriolen zijn niet aanwezig in de centrosomen van andere eukaryotische soorten, zoals planten en de meeste schimmels.
G2-fase (Tweede Gap)
In de G2-fase vult de cel zijn energievoorraden aan en synthetiseert hij eiwitten die nodig zijn voor de chromosoommanipulatie. Sommige celorganellen worden gedupliceerd, en het cytoskelet wordt ontmanteld om middelen te verschaffen voor de mitotische fase. Tijdens G2 kan er sprake zijn van extra celgroei. De laatste voorbereidingen voor de mitotische fase moeten zijn voltooid voordat de cel de eerste fase van mitose kan ingaan.
De mitotische fase
Om twee dochtercellen te maken, moet de inhoud van de celkern en het cytoplasma worden verdeeld. De mitotische fase is een meerstappenproces waarbij de gedupliceerde chromosomen worden uitgelijnd, gescheiden en naar tegenovergestelde polen van de cel worden verplaatst, waarna de cel in twee nieuwe identieke dochtercellen wordt verdeeld. Het eerste deel van de mitotische fase, mitose, bestaat uit vijf fasen, die de celdeling tot stand brengen (figuur 5). Het tweede deel van de mitotische fase, cytokinese genoemd, is de fysieke scheiding van de cytoplasmatische componenten in twee dochtercellen. Hoewel de fasen van mitose voor de meeste eukaryoten vergelijkbaar zijn, is het proces van cytokinese heel anders voor eukaryoten die celwanden hebben, zoals plantencellen.
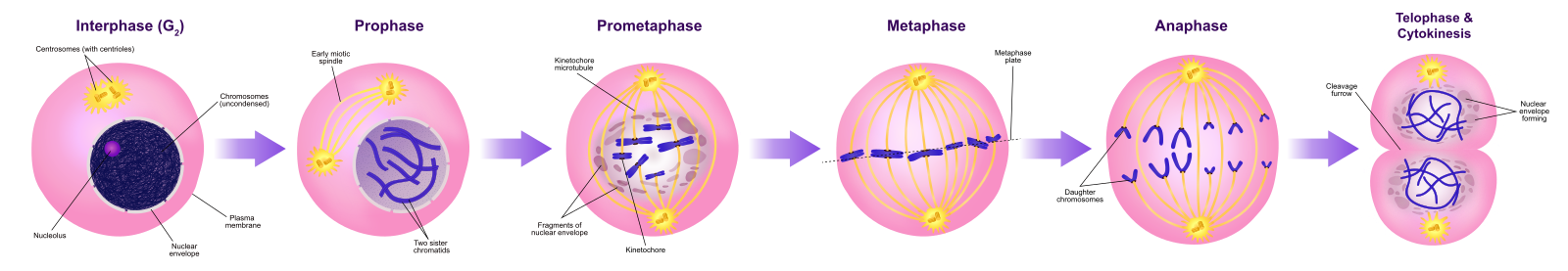
Prophase
Tijdens de prophase, de “eerste fase”, begint het kernomhulsel uiteen te vallen in kleine blaasjes, en de membraneuze organellen (zoals het Golgi-apparaat en het endoplasmatisch reticulum) fragmenteren en verspreiden zich naar de randen van de cel. De nucleolus verdwijnt. De centrosomen beginnen zich te verplaatsen naar de tegenoverliggende polen van de cel. Microtubuli die de mitotische spindel zullen vormen, strekken zich uit tussen de centrosomen en duwen ze verder uit elkaar naarmate de microtubulevezels langer worden. De zusterchromatiden beginnen zich nauwer te omspannen met behulp van condensine-eiwitten en worden zichtbaar onder een lichtmicroscoop.
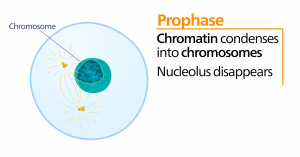
Prometafase
Tijdens de prometafase, de “eerste veranderingsfase”, gaan veel processen die in de profase zijn begonnen, verder. De restanten van de nucleaire envelop fragmenteren. De mitotische spindel ontwikkelt zich verder naarmate meer microtubuli zich verzamelen en uitstrekken over de lengte van het voormalige kerngebied. Chromosomen worden meer gecondenseerd en discreet. Elk zusterchromatide ontwikkelt een eiwitstructuur die kinetochore wordt genoemd in het centromeergebied.
De eiwitten van de kinetochore trekken mitotische spindelmicrotubuli aan en binden deze. Terwijl de spindelmicrotubuli zich vanuit de centrosomen uitstrekken, komen sommige van deze microtubuli in contact met de kinetochores en binden zich er stevig aan vast. Zodra een mitotische vezel zich aan een chromosoom hecht, wordt het chromosoom georiënteerd totdat de kinetochores van de zusterchromatiden naar de tegenovergestelde polen wijzen. Uiteindelijk zullen alle zusterchromatiden via hun kinetochores vastzitten aan microtubuli van tegenover elkaar liggende polen. Spindelmicrotubuli die niet in de chromosomen grijpen, worden polaire microtubuli genoemd. Deze microtubuli overlappen elkaar halverwege tussen de twee polen en dragen bij tot de celverlenging. Astrale microtubuli bevinden zich in de buurt van de polen, helpen bij de oriëntatie van de spindel en zijn nodig voor de regulering van de mitose.

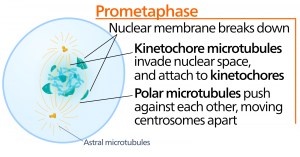
Metafase
Tijdens de metafase, de ‘overgangsfase’, liggen alle chromosomen op één lijn in een vlak dat de metafaseplaat of het equatoriale vlak wordt genoemd, halverwege tussen de twee polen van de cel. De zusterchromatiden zijn nog stevig aan elkaar bevestigd door cohesine-eiwitten. Op dit moment zijn de chromosomen maximaal gecondenseerd.
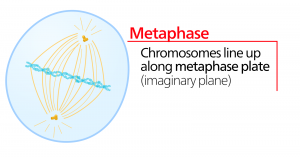
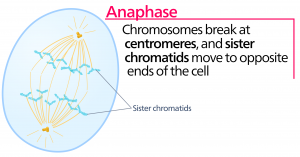
Anafase
Tijdens de anafase, de “opwaartse fase”, worden de cohesine-eiwitten afgebroken en worden de zusterchromatiden bij het centromeer gescheiden. Elk chromatide, dat nu een chromosoom wordt genoemd, wordt snel naar het centrosoom getrokken, waaraan zijn microtubule is bevestigd. De cel wordt zichtbaar langwerpig (ovaalvormig) als de polaire microtubuli tegen elkaar schuiven op de metafaseplaat waar ze elkaar overlappen.
Telofase
Tijdens de telofase, de “afstandsfase”, bereiken de chromosomen de tegenoverliggende polen en beginnen ze te decondenseren (ontrafelen), waarbij ze ontspannen in een chromatineconfiguratie. De mitotische spindels worden gedepolymeriseerd tot tubulinemonomeren die zullen worden gebruikt om cytoskeletcomponenten voor elke dochtercel samen te stellen. Rond de chromosomen vormen zich nucleaire omhulsels, en binnen het nucleaire gebied verschijnen nucleosomen.
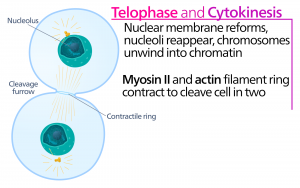
Cytokinesis
Cytokinesis, of “celbeweging”, is het tweede hoofdstadium van de mitotische fase waarin de celdeling wordt voltooid via de fysieke scheiding van de cytoplasmatische componenten in twee dochtercellen. De deling is pas voltooid wanneer de celbestanddelen zijn gesplitst en volledig zijn gescheiden in de twee dochtercellen. Hoewel de stadia van mitose voor de meeste eukaryoten vergelijkbaar zijn, is het proces van cytokinese heel anders voor eukaryoten die celwanden hebben, zoals plantencellen.
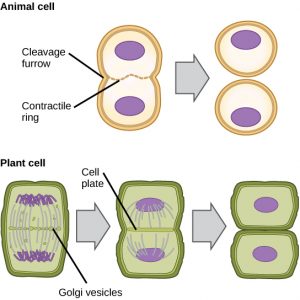
In cellen zoals dierlijke cellen die geen celwanden hebben, volgt cytokinese op het begin van de anafase. Een samentrekkende ring, bestaande uit actinefilamenten, vormt zich net binnen het plasmamembraan op de voormalige metafaseplaat (figuur 12). De actinefilamenten trekken de evenaar van de cel naar binnen, waardoor een kloof ontstaat. Deze kloof, of “barst”, wordt de splijtingsgroef genoemd. De groef wordt dieper naarmate de actinering samentrekt, en uiteindelijk wordt het membraan in tweeën gespleten.
In plantencellen moet zich een nieuwe celwand vormen tussen de dochtercellen. Tijdens de interfase accumuleert het Golgi-apparaat enzymen, structurele eiwitten en glucosemoleculen voordat het in blaasjes uiteenvalt en zich door de delende cel verspreidt (figuur 12). Tijdens de telofase worden deze Golgi-blaasjes getransporteerd op microtubuli om een phragmoplast (een vesiculaire structuur) te vormen op de metafaseplaat. Daar versmelten de blaasjes en smelten samen vanuit het centrum naar de celwanden toe; deze structuur wordt een celplaat genoemd. Naarmate meer blaasjes samensmelten, wordt de celplaat groter en versmelt zij met de celwanden aan de rand van de cel. Enzymen gebruiken de glucose die zich tussen de membraanlagen heeft opgehoopt om een nieuwe celwand te bouwen. De Golgi-membranen worden delen van het plasmamembraan aan weerszijden van de nieuwe celwand.
Samenvatting van Mitose en Cytokinese
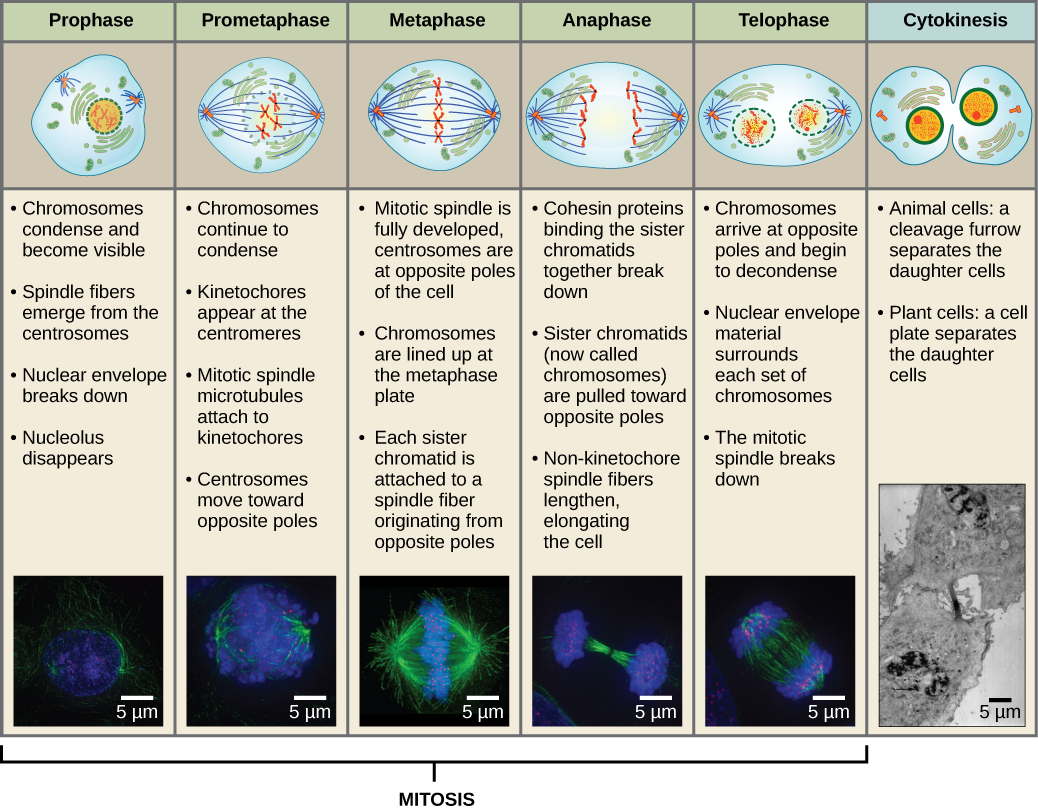
Figuur 13 Mitose is verdeeld in vijf stadia – de prometafase, de metafase, de anafase en de telofase. De foto’s onderaan zijn gemaakt met fluorescentiemicroscopie van cellen die kunstmatig zijn gekleurd met fluorescerende kleurstoffen: blauwe fluorescentie geeft DNA (chromosomen) aan en groene fluorescentie microtubuli (spindelapparaat). (credit “mitose tekeningen”: bewerking van werk van Mariana Ruiz Villareal; credit “microfoto’s”: bewerking van werk van Roy van Heesbeen; credit “cytokinesis microfoto”: Wadsworth Center/New York State Department of Health; schaalbalkgegevens van Matt Russell)
G0-fase
Niet alle cellen houden zich aan het klassieke celcycluspatroon waarin een nieuw gevormde dochtercel onmiddellijk in de interfase komt, op de voet gevolgd door de mitotische fase. Cellen in de G0-fase bereiden zich niet actief voor op deling. De cel bevindt zich in een ruststadium (inactief), omdat hij de celcyclus heeft verlaten. Sommige cellen komen tijdelijk in G0, totdat een extern signaal het begin van G1 inluidt. Andere cellen die zich nooit of zelden delen, zoals rijpe hartspier- en zenuwcellen, blijven permanent in G0).