Introduction
Charophytes of basale streptophytes; (Becker en Marin, 2009; Leliaert et al., 2012) vormen een gevarieerd taxonomisch assortiment van extante zoetwater- en terrestrische groene algen die een breed scala van eencellige, draadvormige en parenchymateuze vormen vertonen (Graham, 1993; Lewis en McCourt, 2004). Een voorouderlijke lijn van charofyten ontstond op het land en koloniseerde dit 450-500 miljoen jaar geleden. Deze organismen pasten zich aan de terrestrische omstandigheden aan, konden overleven en zich voortplanten wanneer ze volledig aan de atmosfeer werden blootgesteld, en sommige leden evolueerden uiteindelijk tot landplanten (Zhong et al., 2013; Delwiche and Cooper, 2015). Deze “terrestrische” ontwikkeling van groene planten was een belangrijke biologische gebeurtenis die de biogeochemie en de natuurlijke geschiedenis van de planeet voor altijd heeft veranderd. De daaropvolgende proliferatie van landplanten veranderde de atmosferische en verder veranderde bodemomstandigheden en maakte de opkomst van andere diverse levensvormen op het land mogelijk. De evolutie van landplanten heeft ook de geschiedenis van de mensheid ingrijpend veranderd, met name door de innovatie van de landbouw en het daaruit voortvloeiende ontstaan van de moderne menselijke beschaving. Mede door hun evolutionaire betekenis hebben charofyten de afgelopen decennia veel aandacht gekregen van plantenbiologen (Pickett-Heaps and Marchant, 1972; Pickett-Heaps, 1975; Mattox and Stewart, 1984; Becker and Marin, 2009; Harholt et al., 2016). Naarmate er echter meer nieuwe gegevens zijn verzameld over de biologie van deze algen, zijn ze belangrijke modellen geworden voor het begrijpen van basisfenomenen in de biochemie, celbiologie, ontwikkelingsbiologie, ecologie en in toenemende mate ook moleculaire biologie (Delwiche en Cooper, 2015; Lemieux et al., 2016). De Frontiers in Plant Science serie over “Charophytes” illustreert het belang van deze organismen in verschillende specifieke gebieden van plantenbiologisch onderzoek. Deze mini-review belicht de eigenschappen van charofyten als modelorganismen in diverse onderzoeksgebieden. Het zal hopelijk ook een aanmoediging zijn voor een nieuwe generatie wetenschappers om het gebruik van deze algen in fundamenteel onderzoek uit te breiden en screening van andere taxa te initiëren, die op hun beurt kunnen leiden tot de identificatie en het gebruik van nieuwe modelcharofyten.
De “Line-Up” van Charofyten
In de jaren zeventig en tachtig van de vorige eeuw zijn ultrastructurele (bijv, celdelingsmechanisme, substructuur van het flagellaire apparaat) en biochemische (bv. enzymprofielen) gegevens de belangrijkste criteria voor de opname van een groene alg in de charofytenlijn (Mattox en Stewart, 1984; Lewis en McCourt, 2004; Leliaert et al., 2012). Vanaf de jaren negentig tot heden hebben studies gericht op moleculaire analyses van chloroplast- en nucleaire genomen en transcriptomen eerdere taxonomische en fylogenetische schema’s herbevestigd en verfijnd (Timme et al., 2012; Delwiche en Cooper, 2015). Dit is verder aangevuld met gedetailleerde biochemische, immunologische en celbiologisch gebaseerde analyses van charofyten celwanden en hormoon biosynthese en signaalwegen (Popper en Fry, 2003; Popper, 2008; Sørensen et al., 2010, 2011, 2012; Zhang en van Duijn, 2014; Ju et al., 2015; O’Rourke et al., 2015). Bestaande charofyten vertonen een relatief laag percentage aan diversiteit in vergelijking met andere groene algentaxa en omvatten 13 families en 122 genera (Becker en Marin, 2009; Leliaert et al., 2012). De huidige fylogenetische opvatting plaatst de charofyten in zes klassen (Figuur 1; Delwiche en Cooper, 2015). De basale klasse, de Mesostigmatophyceae, wordt vertegenwoordigd door één enkel bekend genus, Mesostigma. Deze alg is een eencellige biflagellate met een unieke asymmetrische vorm (d.w.z. als een afgeplatte reddingsboot) en is bedekt met lagen van sierlijk ontworpen schubben (Manton en Ettl, 1965; Becker et al., 1991; Domozych et al., 1991, 1992). De tweede klasse, de Chlorokybophyceae, bestaat eveneens uit één enkele bekende soort, Chlorokybus atmophyticus. Deze zeldzame alg vormt een sarcinoïde pakket cellen omgeven door een dikke gelachtige bedekking (Rogers et al., 1980). Opgemerkt zij dat alternatieve fylogenieën Mesostigma en Chlorokybus als zusterlijnen plaatsen die samen zuster zijn van alle andere Streptofyten (Rodríguez-Ezpeleta et al., 2007). In de fylogenie van de zes klassen bestaat de derde klasse, de Klebsormidiophyceae, uit drie geslachten die eenvoudige onvertakte filamenten vormen (Sluiman et al., 2008) die vaak deel uitmaken van “biologische korsten” die groeien op oppervlakken van diverse terrestrische habitats. Verscheidene soorten van deze klasse zijn zelfs bestand tegen aanzienlijke uitdroging wanneer ze deel uitmaken van woestijnkorsten (Mikhailyuk et al., 2008, 2014; Holzinger en Karsten, 2013); andere zijn tolerant voor de uitdrogingstress die gepaard gaat met lage temperatuuromgevingen (Stamenkovic et al., 2014; Herburger en Holzinger, 2015). Deze eerste drie klassen vormen de “vroege divergente” charofyten. De “laat divergente” lineage omvat ook drie klassen. De Charophyceae, of steenmossen, worden algemeen aangetroffen in zoetwaterecosystemen en bezitten macroscopische meercellige thalli die bestaan uit aggregaties van vertakte filamenten van uitzonderlijk langwerpige cellen (Lewis en McCourt, 2004). Leden van deze groep vertonen een op oogamie gebaseerde geslachtelijke voortplanting met beweeglijk sperma en niet-beweeglijke eitjes, beide geproduceerd in meercellige gametangia. De Coleochaetophyceae bestaan uit taxa die meercellige thalli hebben die bestaan uit vertakte filamenteuze of parenchymachtige thalli (Graham, 1993; Delwiche et al., 2002). Deze organismen vertonen ook een duidelijke op oogamie gebaseerde geslachtelijke voortplanting. De Coleochaetophyceae worden meestal aangetroffen op het grensvlak van zoetwater en terrestrische habitats, vaak als epifyten. De grootste en meest diverse groep charofyten ten slotte, de Zygnematophyceae, bestaat uit eencelligen en onvertakte filamenten (Gontcharov et al., 2003; Guiry, 2013). Het onderscheidende kenmerk van deze groep is de aanwezigheid van conjugatie-gebaseerde geslachtelijke voortplanting die complexe cel-celsignalering en adhesie vereist (Abe et al., 2016). De zygnematofyten algen zijn veel voorkomende bewoners van zoetwater habitats, soms voorkomend in spectaculaire efemere bloei, maar ook in vochtige terrestrische substraten.

FIGUUR 1. Uitheemse Charofyten. Uitheemse Charofyten worden onderverdeeld in vroege en late divergerende taxa. Tot de vroeg divergerende taxa behoren de Mesostigmatophyceae, de Chlorokybophyceae en de Klebsormidiophyceae en tot de laat divergerende taxa behoren de Charophyceae, de Coleochaetophyceae en de Zygnematophyceae. Vertegenwoordigers van elke groep zijn Mesostigma (Mesostigmatophyceae), Chlorokybus atmophyticus (Chlorokybophyceae), Klebsormidium flaccidum (Klebsormidiophyceae), Chara (Charophyceae), Coleochaete scutata (Coleochaetophyceae), en Micrasterias (Zygnematophyceae).
Charophyten als modelorganismen
Verschillende charophyten zijn op grote schaal gebruikt als modelorganismen bij de studie van fundamentele biologische processen. Hun kleine en eenvoudige thalli (d.w.z. in vergelijking met landplanten) en hun gemakkelijke experimentele manipulatie zijn slechts twee van hun attributen die hen aantrekkelijke modelorganismen maken. Recent bewijs heeft ook aangetoond dat veel charofyten verschillende opmerkelijk vergelijkbare kenmerken hebben als die van landplanten, waaronder de aanwezigheid van biosynthetische routes voor veel groeiregulatoren (Figuur 2; Boot et al., 2012; Hori et al., 2014; Wang et al., 2014, 2015; Holzinger en Becker, 2015; Ju et al., 2015) en meerdere celwandpolymeren (Popper, 2008; Sørensen et al., 2010, 2011, 2012; Mikkelsen et al., 2014). Deze twee kenmerken hebben charofyten doeltreffend gemaakt in studiegebieden zoals moleculaire ontwikkeling van planten en stressfysiologie. Hoewel veel charofyten zijn gebruikt in een breed scala van biologische studies, vallen de volgende taxa het meest op door hun uitgebreide gebruik in meerdere studiegebieden als model:
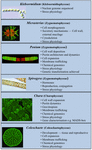
FIGUUR 2. Huidig model Charophyten. Verscheidene Charofyten, waaronder Klebsormidium, Micrasterias, Penium, Spirogyra, Chara en Coleochaete, worden als model gebruikt; elk heeft specifieke eigenschappen waardoor het een bijzonder geschikt model is voor specifieke fysiologische en biochemische processen.
Desmiden: de symmetrische modellen
Onlangs is aangetoond dat de Zygnematophyceae hoogstwaarschijnlijk de meest nabije levende voorouders van landplanten zijn (Wodniok et al, 2011; Delwiche en Cooper, 2015). Eencellige leden van de inclusieve placodermid desmid groep zijn belangrijke modellen geworden voor het ophelderen van veel fundamentele principes van plantencelbiologie en ontwikkeling. Grote celgrootte, opmerkelijke symmetrie/vorm, aparte celwandarchitectuur en uitgebreide endomembraan/cytoskeletale systemen zijn slechts enkele van de kenmerken die hen tot uitstekende celsystemen maken voor het bestuderen van celfysiologie en ontwikkeling. De recente totstandkoming van stabiele getransformeerde lijnen en binnenkort te sequencen genomen/transcriptomen van geselecteerde sieralgsoorten hebben hun waarde voor botanische studies nog vergroot. Twee taxa, Micrasterias en Penium, zijn naar voren gekomen als de meest goed bestudeerde van de sieralgen.
Micrasterias is de afgelopen 50 jaar de meest populaire sieralg geweest voor celbiologisch onderzoek vanwege zijn unieke kenmerken (Meindl, 1993; Lutz-Meindl, 2016). De Micrasterias-cel vertoont een bilaterale symmetrie die vaak wordt benadrukt door een spectaculair ontlede (d.w.z., meerlobbige) periferie. Nieuwe dochter semicellen, geproduceerd door celdeling, hebben deze complexe morfologie niet, maar zijn eerder bolvormig. Een complex post-cytokinetisch ontwikkelingsprogramma dat gebruik maakt van meerdere reeksen van sterk gecoördineerde subcellulaire mechanismen genereert vervolgens het meerlobbige fenotype tijdens de expansie van de dochter semicel. Dit programma is ook verantwoordelijk voor de afzetting van polymeren voor de productie van zowel een primaire als een secundaire celwand. Deze gebeurtenissen zijn gecentreerd rond een groot netwerk van Golgi-lichaampjes en geassocieerde vesikels die een complex secretorisch mechanisme opleveren. Dit proces is op zijn beurt gericht op specifieke plaatsen op het celoppervlak die een multipolaire celexpansie genereren en de gelijktijdige secretie van celwandmacroomoleculen en extracellulaire slijmstoffen (Kim et al., 1996; Lutz-Meindl en Brosch-Salomon, 2000; Oertel et al., 2004; Aichinger en Lutz-Meindl, 2005). Het afleveren van secretorische componenten aan precieze celoppervlak loci vereist een uitgebreid actinomyosine-gebaseerd cytomotiel systeem en wordt gecontroleerd door verschillende signaaltransductie cascades (Meindl et al., 1994; Oertel et al., 2003). Bovendien is de celmorfogenese zeer gevoelig voor externe stress (bv. oxidatieve stress, zoutgehalte, zware metalen) die leidt tot grote veranderingen in de celdifferentiatie (Darehshouri et al., 2008; Affenzeller et al., 2009; Andosch et al., 2012). Micrasterias is een uitstekend organisme voor het analyseren van deze subcellulaire kenmerken omdat het gemakkelijk te onderhouden en te manipuleren is in het laboratorium en het leent zich vooral goed voor het verkrijgen van hoge resolutie beeldvorming met behulp van licht- en elektronenmicroscopie, waaronder immunocytochemie, Focused Ion Beam-Scanning Electron Microscopy (FIB-SEM) en Electron Energy Loss Spectroscopy (EELS) beeldvorming (Lutz-Meindl, 2007; Eder en Lutz-Meindl, 2008, 2010; Wanner et al., 2013; Lutz-Meindl et al., 2015). Bovendien hebben initiële moleculaire analyses, waaronder de productie van getransformeerde cellijnen, het gebruik van Micrasterias verder verbeterd, vooral in de moleculaire dynamiek van celwandverwerking (Vannerum et al., 2010, 2011, 2012).
In het afgelopen decennium is ook de sieralg, Penium margaritaceum, een waardevol modelorganisme geworden (Domozych et al., 2009, 2014; Sørensen et al., 2014; Rydahl et al., 2015; Worden et al., 2015). In tegenstelling tot Micrasterias heeft Penium een eenvoudige cilindrische vorm, bezit het slechts een primaire celwand en zet het tijdens de expansie wandpolymeren af op twee specifieke loci van het celoppervlak (Domozych et al., 2011). Deze relatieve eenvoud is zeer aantrekkelijk voor het ophelderen van fundamentele principes van plantencelontwikkeling, waaronder celwandontwikkeling, celexpansie en secretiedynamiek (Domozych et al., 2005, 2014; Ochs et al., 2014). Een van de belangrijkste eigenschappen is dat het wandpolymeren heeft die vergelijkbaar zijn met die in veel landplanten (bv. cellulose, pectinen, hemicellulosen) en dat deze polymeren kunnen worden gelabeld met monoklonale antilichamen. Na het labelen van levende cellen kunnen deze opnieuw in cultuur worden gebracht, waar de daaropvolgende celexpansie en wandafzetting kunnen worden gevolgd (Domozych et al., 2009; Rydahl et al., 2015). Penium is ook gemakkelijk te onderhouden in het laboratorium en zijn snelle groeisnelheid onder nauwkeurig gecontroleerde omstandigheden maakt het een uitstekend specimen voor grootschalige gelijktijdige microarray screenings van vele chemische agentia (door groei in multi-well platen) en voor de beoordeling van hun specifieke effecten op expansie/wandontwikkeling (Worden et al., 2015). Dit helpt aanzienlijk bij het onthullen van de rol van specifieke subcellulaire componenten en processen in de expansie/differentiatie. Onlangs heeft de isolatie van stabiele getransformeerde lijnen (Sørensen et al., 2014) het potentieel van deze alg in toekomstige plantencelstudies verder versterkt.
Chara en Nitella: Cellular Giants for Plant Physiology
Soorten van Chara en Nitella (Charophyceae) zijn lang gebruikt als specimens voor een verscheidenheid aan biologische onderzoeken, vooral die welke te maken hebben met cellulaire dynamiek, expansie en cytoplasmatische streaming (Green, 1954; Probine en Preston, 1962). Hun macroscopische thalli onderscheiden zich door knopen, waar vertakkingen en gametangia ontstaan, en internodale gebieden die bestaan uit uitzonderlijk langwerpige cellen. Internodale cellen vertonen een duidelijk waarneembare gelaagdheid van subcellulaire componenten waarbij spiraalvormig georiënteerde chloroplasten een stationaire cortex definiëren die een endoplasma omgeeft waar de snelst geregistreerde actinomyosine-gegenereerde cytoplasmastroom plaatsvindt. Het cytoplasma bevat diverse endomembraancomponenten die deelnemen aan dynamische netwerken voor membraantransport (bv. endocytose en exocytose) die worden gecontroleerd door complexe signaaltransductiecascades (Foissner en Wastenys, 2000, 2012, 2014; Sommer et al., 2015; Foissner et al., 2016). De internodale cellen bezitten celwanden die rijk zijn aan cellulose en de pectine, homogalacturonan (HG). Turgor is de belangrijkste drijvende kracht voor expansie en pectine is hoogstwaarschijnlijk de dragende component die de wandexpansie controleert. Cyclische, niet-enzymatische modulatie en gelijktijdige calciumcomplexering van de HG is het mechanisme dat verantwoordelijk is voor het controleren van wandmodulaties die leiden tot celexpansie (Proseus en Boyer, 2007, 2008, 2012; Boyer, 2016). Een van de technische voordelen van Chara en Nitella bij celonderzoek is dat internodale cellen individueel kunnen worden verwijderd; endoplasma en celwanden kunnen worden “ontleed” en vervolgens worden gebruikt als acellulaire systemen voor experimentele manipulatie.
Andere thalluscomponenten zoals rhizoïden hebben een georganiseerde polaire organisatie van het cytoplasma. Deze cellen bevatten sedimenteerbare, mineraalrijke statolieten op de apex die functioneren in de waarneming van de zwaartekracht (Hodick et al., 1998; Braun, 2002; Braun and Limbach, 2006). Deze eigenschap heeft Chara tot de eerste charofyt in de ruimte gemaakt. Tijdens parabolische vluchten van de TEXUS raket werd aangetoond dat de statolieten trekkrachten uitoefenen op actine filamenten. Een evenwicht van krachten (nl. de zwaartekracht en de tegenwerkende kracht van actine filamenten) is verantwoordelijk voor de juiste positionering van de statolieten in het rhizoid, die op hun beurt de mogelijkheid garanderen om te reageren op de zwaartekrachtsvector (Braun en Limbach, 2006). Het gebruik van laser pincetten en langzaam roterende centrifuge microscopie heeft aangetoond dat statoliet sedimentatie niet voldoende is om gravitropische buiging van de rhizoïd tip te veroorzaken. De mineraalrijke statolieten moeten zich eerder op specifieke delen van het plasmamembraan afzetten om gravitropische morfologische effecten te bewerkstelligen (Braun, 2002). Chara is ook gebruikt om andere fenomenen te bestuderen, waaronder elektrofysiologie en de rol en beweging van hormonen (Belby, 2016).
Coleochaete voor ontwikkelingsstudies en patroonontwikkeling
De vorming van een meercellig thallus (d.w.z. weefsel, orgaan) van een plant vereist nauwkeurige, ruimtelijk gereguleerde celdelingsvlakken die worden gecontroleerd door meerdere sets van genen die op specifieke punten tijdens de ontwikkeling tot expressie komen (Besson en Dumais, 2011; Umen, 2014). Geleid in zowel 3-dimensionale ruimte als tijd, moduleert de thallus morfogenese ook in reactie op externe stressfactoren. In planten worden deze ontwikkelingsprogramma’s nog complexer gemaakt door de aanwezigheid van celwanden die geen celmigratie of weefselflexibiliteit toelaten. De opheldering van de specifieke gebeurtenissen die centraal staan bij de manifestatie van thallus vorm en grootte is vaak moeilijk te ontcijferen bij landplanten waar de resolutie van specifiek celgedrag vaak slecht is wanneer ingebed in zeer complexe en uitgestrekte weefsels/organs. Het gebruik van organismen met eenvoudiger thallusontwerpen is voordelig gebleken voor het begrijpen van fundamentele ontwikkelingsfenomenen en het charofytengeslacht Coleochaete is zo’n voorbeeld. Bepaalde soorten (bv. C. orbicularis, C. scutatum) produceren een parenchymateus discoïde thallus dat naar buiten groeit door een combinatie van anti- of peri-clinale celdelingen en de daaropvolgende expansie van de buitenste cellen (Brown et al., 1994; Cook, 2004). Het delingsvlak van elke cel volgt eenvoudige regels die gebaseerd zijn op celgrootte, celvorm en geografische locatie in het thallus (Dupuy et al., 2010; Besson en Dumais, 2011). Deze eigenschap maakt de constructie van wiskundige modellen mogelijk die vervolgens kunnen worden gebruikt om de meercellige morfogenese te interpreteren (Domozych en Domozych, 2014; Umen, 2014) en kritisch inzicht opleveren in de evolutie, biomechanica en fysiologie van organen en hele organismen. Bovendien zijn sommige Coleochaete-soorten uitgebreid bestudeerd om de structurele en functionele modulaties te bepalen die zich voordoen in een thallus onder uitdroging (Graham et al., 2012), d.w.z. belangrijke kenmerken om de evolutie van vroege landplanten en de groeidynamiek van planten tijdens droogtes te begrijpen.
Inzichten uit genoom- en genstudies
Gelijkaardig aan vele andere technologieën hebben moleculaire instrumenten de mogelijkheid om nieuwe informatie te verschaffen die van belang is voor vele onderzoeksgebieden, waaronder fysiologie en ontwikkeling. Zo zijn leden van de MADS-box genenfamilie geïsoleerd en gekarakteriseerd van drie charofyten, Chara globularis, Coleochaete scutata, en Closterium peracerosum-strigosum-littorale (Tanabe et al., 2005). Het expressiepatroon van de MADS-box genen in de charofyten suggereert dat zij een rol spelen in haploïde ontwikkeling en voortplanting (Tanabe et al., 2005). Er wordt aangenomen dat ze tijdens de evolutie van landplanten zijn gerekruteerd in de diploïde generatie, en dat hun rol in de ontwikkeling verder is gediversifieerd (Tanabe et al., 2005). Tot op heden is Klebsormidium flaccidum de enige charofyt waarvoor een bijna volledig nucleair genoom beschikbaar is (Hori et al., 2014); hoewel chloroplast- en mitochondriale genomen zijn gesequenced voor vertegenwoordigers van elk van de zes klassen van charofyten en EST’s (expressed sequence tags) beschikbaar zijn voor sommige charofyten (Delwiche, 2016). Het Klebsormidium-genoom heeft al de aanwezigheid onthuld van genen voor de synthese van verschillende plantenhormonen en signaalintermediairs, mechanismen voor bescherming tegen hoge lichtintensiteit (Hori et al., 2014), en de aanwezigheid van Groep IIb WRKY-transcriptiefactoren waarvan eerder werd gedacht dat ze voor het eerst in mossen voorkwamen (Rinerson et al., 2015). Verschillende charofyten genoom sequencing projecten zijn momenteel aan de gang (Delwiche, 2016) en zullen meer gedetailleerde vergelijkende analyses mogelijk maken met inbegrip van een groter aantal charofyten en mogelijk nieuw inzicht geven in de evolutionaire relaties tussen soorten en genen. De unieke positie van charofyten als de vroegst divergerende groep van Streptofyten maakt ze tot een bijzonder krachtig instrument voor evolutionaire-ontwikkelingsstudies en opkomende technologieën, zoals het recente bereiken van stabiele transformatie in Penium (Sørensen et al, 2014) zullen het mogelijk maken de functie van specifieke genen te onderzoeken.
Toekomstige toepassingen van Charofyten als modelorganismen
Een cruciale volgende stap in het charofytenonderzoek is de analyse van de genomen van inclusieve taxa en vergelijkende studies met die van landplanten. Niet alleen zal dit werk belangrijke inzichten opleveren in de aanpassingsmechanismen die charofyten hebben ontwikkeld bij hun opkomst op, en succesvolle kolonisatie van, terrestrische habitats, maar het zal ook het gebruik van de huidige modelorganismen in fundamenteel plantenonderzoek verder verfijnen en uitbreiden. Hoewel moleculaire studies vele onderzoeksgebieden zullen ondersteunen, zijn er twee gebieden die onmiddellijk van nut zullen zijn: celwandbiologie en de dynamiek van groeiregulatoren in groei en ontwikkeling. Op het eerste gebied kunnen de charofyten opheldering verschaffen over vele fundamentele, maar nog niet goed opgeloste verschijnselen, zoals de biosynthese van pectine en celwandeiwitten, de gecontroleerde secretie, afzetting en modulatie na de secretie van wandpolymeren en de specifieke, interactieve netwerken voor de handel in membranen in plantencellen. Wat dit laatste betreft, zouden charofytenmodellen helpen bij het ontcijferen van de moleculaire signalering, de celbiologie en de ontwikkelingsdynamiek van groei en ontwikkeling in verband met hormonen als ethyleen en auxine. Recente ecofysiologische studies met charofyten bieden ook een potentiële bonanza aan kritische gegevens om te bepalen hoe plantencellen zich aanpassen aan stress, waaronder uitdroging en zouttolerantie (Holzinger en Karsten, 2013; Pichrtová et al., 2014; Herburger et al., 2015; Holzinger en Becker, 2015; Holzinger en Pichrtová, 2016; Kondo et al., 2016). De toekomst van charofytenonderzoek is inderdaad zeer helder en zal bijgevolg een zegen worden voor de hele plantenbiologie.
Author Contributions
Alle vermelde auteurs, hebben een substantiële, directe en intellectuele bijdrage geleverd aan het werk, en het goedgekeurd voor publicatie.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek is uitgevoerd in de afwezigheid van commerciële of financiële relaties die kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgment
Dit werk werd ondersteund door NSF (USA) MCB collaborative grants 1517345 en 1517546.
Belby, M. J. (2016). Multi-scale characean experimenteel systeem: van elektrofysiologie van membraantransporters naar cel-naar-cel connectiviteit, cytoplasmatische streaming en auxine metabolisme. Front. Plant Sci. 7:1052. doi: 10.3389/fpls.2016.01052
CrossRef Full Text | Google Scholar
Braun, M. (2002). Gravity perception requires statoliths settled on specific plasma membrane areas in characean rhizoids and protonemata. Protoplasma 219, 150-159. doi: 10.1007/s007090200016
CrossRef Full Text | Google Scholar
Braun, M., and Limbach, C. (2006). Rhizoids and protonemata of characean algae: model cells for research on polarized growth and plant gravity sensing. Protoplasma 229, 133-142. doi: 10.1007/s00709-006-0208-9
CrossRef Full Text | Google Scholar
Graham, L. E. (1993). Oorsprong van landplanten. New York, NY: John Wiley & Sons, 287.
Google Scholar
Herburger, K., and Holzinger, A. (2015). Localization and quantification of callose in the Streptophyte green algae Zygnema and Klebsormidium: correlation with desiccation tolerance. Plant Cell Physiol. 56, 2259-2270. doi: 10.1093/pcp/pcv139
CrossRef Full Text | Google Scholar
Lemieux, C., Otis, C., and Turmel, M. (2016). Vergelijkende chloroplast genoomanalyses van streptofyte groenwieren leggen belangrijke structurele veranderingen bloot in de Klebsormidiophyceae, Coleochaetophyceae en Zygnematophyceae. Front. Plant Sci. 7:697. doi: 10.3389/fpls.2016.00697
CrossRef Full Text | Google Scholar
Lutz-Meindl, U. (2007). Gebruik van energiefilterende transmissie-elektronenmicroscopie voor beeldvorming en elementanalyse in plantaardige organismen. Micron 38, 181-196. doi: 10.1016/j.micron.2006.03.017
CrossRef Full Text | Google Scholar
Meindl, U. (1993). Micrasterias cellen als modelsysteem voor onderzoek naar morfogenese. Microbiol. Rev. 57, 415-433.
Google Scholar
Pichrtová, M., Kulichova, J., and Holzinger, A. (2014). Stikstofbeperking en langzame uitdroging induceren uitdrogingstolerantie in conjugerende groenwieren (Zygnematophyceae, Streptophyta) uit polaire habitats. PLoS ONE 9:e113137. doi: 10.1371/journal.pone.0113137
CrossRef Full Text | Google Scholar
Pickett-Heaps, J. D., and Marchant, H. J. (1972). De fylogenie van groene algen: een nieuw voorstel. Cytobios 6, 255-264.
Google Scholar
Stamenkovic, M., Woelken, E., and Hanelt, D. (2014). Ultrastructure of Cosmarium strains (Zygnematophyceae, Streptiophyta) collected from various geographic locations shows species-specific differences both at optimal and stress temperatures. Protoplasma 251, 1491-1509.
Google Scholar