Wprowadzenie
Poprzednia praca z naszego laboratorium (Shalom, 2009) dowodziła, że przetwarzanie neurokognitywne może być podzielone na trzy poziomy: (1) poziom podstawowy, obejmujący pierwotne przetwarzanie poznawcze, emocjonalne i sensomotoryczne. Na przykład, głośny, niespodziewany dźwięk, który jest postrzegany przez pierwotne systemy słuchowo-zmysłowe, może wywołać fizjologiczne reakcje emocjonalne, takie jak strach (szybkie bicie serca, itp.). (2) poziom integracyjny, który łączy wyjście wszystkich pierwotnych procesów z poziomu podstawowego i tworzy globalne, spójne znaczenie, doświadczenie lub zachowanie. Na przykład, mentalne reprezentacje różnych elementów podstawowych, które tworzą reakcję emocjonalną strachu, skutkują świadomym uczuciem strachu. (3) Poziom logiczny, który tworzy abstrakcyjne reguły logiczne (zasady „jeśli – to”) z poziomu podstawowego (np. „Jeśli mam szybkie bicie serca i zimny pot, to mogę się bać”), a także jest zaangażowany w selekcję i hamowanie (np. kontrolowanie popędów emocjonalnych). Ta trzypoziomowa architektura została zastosowana do czterech ogólnych domen psychologicznych: emocji, pamięci, percepcji i motoryki. Shalom (2009) skupił się na drugim, integracyjnym poziomie i jego związku z czterema domenami psychologicznymi. Stwierdził również, że te cztery rodzaje integracji są obsługiwane przez cztery różne podobszary przyśrodkowej kory przedczołowej: Obszar Brodmanna (BA) 11 (percepcja), BA 10 (pamięć), BA 9 (emocje) i BA 8 (motoryka). Wreszcie, przedstawiono dowody na to, że selektywny deficyt w tych obszarach BA i te rodzaje procesów integracyjnych leżą u podstaw niektórych wspólnych deficytów w ASD (zaburzenia ze spektrum autyzmu).
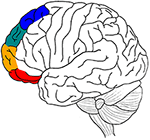
Rysunek 1. Graficzne zestawienie głównych hipotez anatomicznych artykułu. Motor processing in blue, emotion in green, memory in orange, and sensory-perceptual in red.
Perception
Perceptual Selection and Inhibition and Lateral BA 11/47
Istnieją dowody na to, że percepcyjna selekcja i hamowanie są wspierane przez sieci neuronalne angażujące boczną część BA 11/47 w LPFC. W modelu zaproponowanym przez Zsugę i wsp. (2016) sugeruje się, że kora orbitofrontalna (OFC) odgrywa główną rolę w selekcji bodźców wzrokowych zgodnie z wymaganiami zadania.W szczególności sugerują oni, że przyśrodkowa część OFC odgrywa centralną rolę w integracji i kategoryzacji informacji o bodźcach ze środowiska i ich kontekście, podczas gdy boczna część OFC jest zaangażowana w przypisywanie i aktualizację parametrów selekcji zgodnie ze specyficznymi dla zadania wartościami dostępnych bodźców. Na poparcie tego twierdzenia, w jednym z badań Howard i Kahnt (2017) wykazali, że boczna część BA 11 jest zaangażowana w kodowanie wartości bodźców węchowych ukierunkowanych na cel. Prezentowali oni głodnym uczestnikom dwa preferowane zapachy jedzenia i wymagali od nich wyboru, czy chcą wąchać jeden czy drugi. Później uczestnicy otrzymywali do zjedzenia jeden z dwóch pokarmów i ponownie mogli wybierać pomiędzy dwoma zapachami. Zmiana preferencji w kierunku zapachu nienasyconego została odzwierciedlona przez zmianę aktywności w bocznej części BA 11/47. Ponadto wykazano, że boczna część BA 11/47 jest zaangażowana w odrzucanie nieistotnych bodźców (Kaufman i in., 2016).
Wykazano również, że boczna część BA 11/47 jest zaangażowana w utrzymywanie bodźców wzrokowych w pamięci roboczej i kierowanie wyborem wzrokowym. W jednym z badań, Soto i wsp. (2007), porównali dwa warunki primed visual selection, w których prime był pokazywany albo raz, albo dwa razy. Boczna aktywacja BA 11/47 była większa, gdy prym był pokazywany dwukrotnie, jednak redukcję aktywacji stwierdzono, gdy podobne bodźce były pasywnie powtarzane dwukrotnie (bez fazy wyboru wzrokowego, a więc bez znaczenia dla zadania).
Wreszcie, według Price’a (2007), boczne BA 11 jest częścią sensorycznej sieci oczodołowej.
ADHD i wizualna selekcja i hamowanie
Istnieją dowody, że dzieci i dorośli z ADHD mają problemy z hamowaniem nieistotnych bodźców wzrokowych i słuchowych. Na przykład, dorośli z ADHD wykazali więcej efektów interferencyjnych (tj. problemów z hamowaniem nieistotnych bodźców) niż osoby z grupy kontrolnej, gdy poproszono ich o wykonanie zadania przypominania sobie podczas ignorowania hałasu w tle (Pelletier i in., 2016). Co więcej, stwierdzono dodatnią korelację między wykonaniem zadania w warunkach nieistotnego dźwięku a stopniem nasilenia objawów uwagi zgłaszanych przez pacjentów na klinicznej skali objawów. W innym badaniu poproszono dorosłych z ADHD i grupę kontrolną o prowadzenie rozmowy telefonicznej podczas prowadzenia samochodu. Grupa z ADHD wykazała znacząco gorsze umiejętności prowadzenia pojazdu podczas rozmowy telefonicznej niż w warunkach ciszy w porównaniu z grupą kontrolną (Reimer et al., 2010). Inne badania wykazały słabą inhibicję wzrokową u osób z ADHD. Na przykład Forster i wsp. (2014) oraz Forster i Lavie (2016) stwierdzili, że dorośli z ADHD wykonywali zadanie polegające na wyszukiwaniu liter wolniej, gdy na ekranie obecne były nieistotne, ale istotne postacie z kreskówek, w większym stopniu niż osoby z grupy kontrolnej. W pokrewny sposób, zadanie interferencyjne Stroopa wymaga od uczestników zwrócenia uwagi na określony wymiar bodźca wzrokowego przy jednoczesnym ignorowaniu innego. Badania wykazały, że dzieci z ADHD osiągały gorsze wyniki w warunku interferencji Stroopa niż osoby z grupy kontrolnej (Sørensen i in., 2014), oraz że ich wyniki korelowały z objawami nieuwagi i nadpobudliwości (Ikeda i in., 2013).
ADHD, Visual Selection, and Inhibition and Lateral BA 11/47
W dodatku, w kilku badaniach znaleziono specyficzne dowody na atypową aktywację w bocznym BA 11/47 u osób z ADHD wykonujących zadania selekcji wzrokowej/inhibicji. Tsujimoto i wsp. (2013) poprosili dzieci z ADHD i bez ADHD o wykonanie zadania pamięci roboczej z rozproszeniem i bez rozproszenia. Uczestnicy z ADHD wykazywali istotnie gorsze wyniki behawioralne, szczególnie w warunkach rozproszenia uwagi, oraz wykazywali istotnie wyższy poziom aktywacji w bocznym BA 11/47 niż osoby z grupy kontrolnej. W innym badaniu Yasumura i wsp. (2014) testowali dzieci z ADHD i dzieci z grupy kontrolnej zarówno w warunkach interferencji Stroopa, jak i w warunkach odwróconej interferencji Stroopa, i stwierdzili, że w obu warunkach dzieci z ADHD osiągały gorsze wyniki, a wyniki korelowały z aktywnością bocznego BA 47. W innym badaniu, w którym podczas zadania szacowania czasu prezentowano wyraźny, ale nieistotny dystraktor, stwierdzono, że podczas gdy dystraktor pomagał uczestnikom z ADHD w takim samym stopniu jak osobom z grupy kontrolnej, boczna aktywacja BA 11, która towarzyszyła pojawieniu się dystraktora, była znacząco większa (Pretus i in., 2016).
Pamięć
Memory Selection and Inhibition and Lateral BA 10/46
Choć nie ma ogólnej zgody co do tego, że BA10/46 jest zaangażowany w funkcje pamięci epizodycznej, istnieje wiele dowodów na poparcie roli BA 10/46 w selekcji i hamowaniu informacji pamięciowych. Na przykład Kim (2011), analizując 74 badania, w których porównywano zapamiętywanie i zapominanie (tj. pomyślnie zapamiętane elementy vs. dystraktory ocenione jako błędnie występujące), stwierdził, że największa część badań pokrywała się w przedniej części BA 46, w pobliżu jego połączenia z bocznym BA 10. W późniejszym badaniu Kim (2013) przeprowadził metaanalizę 70 badań koncentrujących się na nowych i starych elementach (tj. trafieniach vs. poprawnych odrzuceniach). Wyniki pokazują, że porównania nowe>stare powodowały aktywacje, które rozprzestrzeniały się w środkowej części lewej bocznej części przedczołowej. Jednakże, gdy koncentrujemy się na badaniach pamięci źródłowej, w których uczestnicy muszą odzyskać nie tylko element docelowy, ale także dodatkowy szczegół związany z elementem z fazy uczenia się (tj, gdy wymagania selekcji są wyższe), specyficzną aktywację zaobserwowano w przedniej części BA46, w tym w rostralnej części BA10.
Inne dowody pochodzą z metaanalizy Gilberta i wsp. (2006), w której przeanalizowano 104 badania koncentrujące się na BA 10. Stwierdzono, że większość badań angażujących boczną część BA 10 testowała pamięć roboczą i odzyskiwanie pamięci epizodycznej, podczas gdy badania angażujące przyśrodkową część BA 10 testowały mentalizację (tj, attending to one’s own emotions and mental states or those of other agents).
In addition, Cohen et al. (2014), employed the value directed remembering task, in which participants are presented with word lists in which some words are assigned more value than others, thus encouraging participants to attempt and select higher value items over low value ones during encoding. Wyniki pokazują specyficzną aktywację prawego bocznego BA46/10 i bardzo wyraźną aktywację lewego bocznego BA46/10, gdy pojawiała się wskazówka przewidująca słowo o wysokiej wartości.
ADHD a selekcja i hamowanie pamięci
Istnieje kilka badań, które znalazły dowody na trudności w selekcji i hamowaniu informacji pamięciowych w ADHD. Na przykład, w badaniu Pollak et al. (2007), dorośli z i bez ADHD wykonane trudne zadanie uczenia się listy, w którym musieli zapamiętać i przypomnieć sobie pięć list słów na osiem różnych okazjach. Wyniki pokazują, że uczestnicy z ADHD popełniali więcej podwójnych przywołań i błędów wtargnięcia (przywoływanie słów z niewłaściwej listy), które są przykładem trudności w hamowaniu przywoływania nieistotnych dla zadania elementów. W podobnym badaniu, Soliman i Elfar (2017), poprosili dorosłych z i bez ADHD o nauczenie się ośmiu list, a następnie zaprezentowano im zadanie rozpoznawania. Grupa z ADHD rozpoznała mniej słów poprawnie, wyprodukowała więcej fałszywych pozytywnych odpowiedzi (tj. błędnie wybierając pozycje, które nie zostały nauczone) i była bardziej pewna swoich błędów. W innym badaniu Castel et al. (2011) poprosili dzieci z ADHD i bez ADHD o wykonanie wspomnianego wyżej paradygmatu zapamiętywania ukierunkowanego na wartość. Wyniki pokazują, że dzieci z ADHD przypomniały sobie tyle samo pozycji co dzieci z grupy kontrolnej, jednak były mniej zdolne do wyboru pozycji do zapamiętania.
ADHD, Memory Selection, and Inhibition and Lateral BA 10/46
Wyszukiwanie literatury przyniosło tylko jedno badanie neuroobrazowania oceniające wybór lub hamowanie funkcji pamięciowych w ADHD. W badaniu przeprowadzonym przez Depue et al. (2010), dorośli z i bez ADHD byli uczeni relacji między parami twarz-obraz do momentu opanowania. Później pokazywano im obraz twarzy i proszono, aby albo pomyśleli o powiązanym obrazie, albo nie dopuścili do pojawienia się powiązanego obrazu w świadomości. Wyniki pokazują, że uczestnicy z ADHD aktywowali boczne BA 10 mniej niż kontrolerzy, gdy porównywali warunek „nie myśleć” z warunkiem „myśleć”.
Emotion
Emotion Selection and Inhibition and Lateral BA 9
Istnieje wiele dowodów wspierających związek między regulacją emocji (hamowanie pewnych emocji i wybór innych) a bocznym BA 9. Na przykład w badaniu Hallam i wsp. (2014) uczestnicy oglądali filmy wywołujące emocje i byli zobowiązani albo do stłumienia lub przewartościowania swojej reakcji emocjonalnej, albo po prostu do obejrzenia filmu. Wyniki wykazały, że zarówno przewartościowanie, jak i stłumienie w porównaniu do stanu pasywnego wykazywały aktywację bocznego BA 9. Podobnie w dwóch badaniach porównano przewartościowanie i pasywne oglądanie obrazów u osób z PTSD i w grupie kontrolnej. W obu badaniach stwierdzono, że boczne BA 9 było aktywowane bardziej podczas warunku reappraisal, ale w mniejszym stopniu u osób z PTSD (Xiong i in., 2013; Rabinak i in., 2014).
Dodatkowo do analizy badań fMRI oceniających regulację emocji wykorzystano dwie różne metaanalizy. Buhle i wsp. (2014) stwierdzili, że przewartościowanie obejmujące regulację negatywnego afektu w dół konsekwentnie aktywowało boczną BA 9; Frank i wsp. (2014) stwierdzili, że takiemu przewartościowaniu towarzyszyła większa aktywacja w bocznej BA 9 i zmniejszona aktywacja w amygdali.
ADHD and Emotion Selection and Inhibition
W wielu badaniach stwierdzono deficyty regulacji emocji u dzieci z ADHD. Na przykład, kilka badań pokazuje, że dzieci z ADHD mają trudniejszy czas tłumienia emocji niż ich typowo rozwijających się (TD) rówieśników. Na przykład, w jednym z badań dzieci poddano zadaniu rywalizacji rówieśniczej, w którym połowa została poproszona o ukrycie swoich emocji, a połowa nie. Według ocen dokonanych przez naiwnych sędziów, wyniki pokazały, że dzieci z ADHD były mniej zdolne do maskowania swoich emocji niż ich rówieśnicy (Walcott i Landau, 2004). Ponadto, w badaniu oceniającym wyniki dzieci z ADHD w emocjonalnym zadaniu Stroopa, stwierdzono, że dzieci z ADHD miały trudniejszy czas hamowania reakcji na gniewne i sfrustrowane twarze (Yarmolovsky i in., 2016).
Dodatkowo, w metaanalizie regulacji emocji w ADHD przeprowadzonej przez Graziano i Garcia (2016), znaleźli oni cztery interesujące domeny, a mianowicie: rozpoznawanie (tj, zdolność do przetwarzania i wnioskowania o emocjach innych osób, jak również własnych), reaktywność (tj. próg, intensywność i czas trwania pobudzenia afektywnego), regulacja (tj. skuteczne reagowanie na reaktywność emocjonalną w sposób elastyczny, ułatwiający adaptacyjne funkcjonowanie) oraz empatia (tj. zdolność do doświadczania stanu afektywnego innej osoby i/lub wyrażania troski o jej położenie). Wyniki pokazują, że podczas gdy wszystkie cztery domeny wydają się być nietypowe w populacji ADHD, najbardziej zauważalne braki występują w domenach reakcji i regulacji. Oznacza to, że (szczególnie) dzieci z ADHD mają tendencję do reagowania szybciej, bardziej intensywnie i przez dłuższy czas na sytuacje awersyjne i są mniej zdolne do regulacji tych emocji, nawet jeśli próbują to zrobić.
ADHD, Emotion Selection, and Inhibition and Lateral BA 9
Istnieje kilka badań, które pokazują specyficzną atypowość w bocznej aktywacji BA 9 u uczestników z ADHD wykonujących zadania wymagające regulacji emocji. Na przykład, w badaniu Passarotti i wsp. (2010a), dzieci z zaburzeniem dwubiegunowym, dzieci z ADHD oraz grupa kontrolna dobrana pod względem wieku wykonywały zadanie Stroopa dotyczące walencji emocjonalnej. W zadaniu tym prezentowano pozytywne, neutralne i negatywne słowa, a uczestnicy musieli reagować zgodnie z kolorem słowa, ignorując jego znaczenie. Wyniki pokazują, że grupa dwubiegunowa miała trudności z ignorowaniem słów pozytywnych, podczas gdy grupa ADHD miała trudności z ignorowaniem słów negatywnych. Co ciekawe, boczna aktywacja BA 9 była bardziej wyraźna dla warunku zadania, które zostało wykonane z powodzeniem, tj. dla słów negatywnych w grupie dwubiegunowej i dla słów pozytywnych w grupie ADHD. Ponadto, w zadaniu emocjonalnym opartym na Stroopie, Hwang i wsp. (2015), poprosili dzieci z ADHD i bez ADHD o wykonanie numerycznego zadania Stroopa po pokazaniu im obrazków, które były pozytywne, neutralne lub negatywne. Podczas gdy dane behawioralne nie wykazały znaczących różnic pomiędzy różnymi stanami emocjonalnymi, odkryli oni, że dzieci z ADHD były mniej zdolne do rekrutacji bocznego BA 9 podczas wykonywania zadania, cecha, która również korelowała z nasileniem objawów mierzonym skalą rodzicielską Connera.
Motor
Motor Selection and Inhibition and Lateral BA 8
Istnieje wiele dowodów na to, że hamowanie czynności motorycznych jest wspierane przez sieci neuronalne w bocznym BA 8. Najbardziej powszechną miarą hamowania motorycznego jest zadanie Stop Signal Task (SST), w którym uczestnicy są zobowiązani do naciśnięcia klawisza, gdy prezentowany jest bodziec, ale powstrzymują się od wykonania tego naciśnięcia, jeśli po bodźcu następuje krótki sygnał. Badania pokazują, że hamowanie naciśnięcia klawisza wiąże się z aktywacją w bocznym BA 8 (Matthews i in., 2005; Smith i in., 2013; Hughes i in., 2014). Wykazano również, że boczne BA 8 jest specyficznie aktywne, gdy wybór reakcji motorycznej wymagał uwagi lub wiązał się z konfliktem. Na przykład, Enriquez-Geppert i wsp. (2013), poprosili uczestników o naciśnięcie klawisza, gdy na ekranie pojawił się bodziec, jednak w niektórych próbach klawisz odpowiedzi zmieniał się. Eksperymentatorzy stwierdzili, że boczne BA 8 było specyficznie aktywowane w próbach, w których klawisz odpowiedzi był inny. Podobnie Lenartowicz i wsp. (2011) wykorzystali zadanie typu go/no-go, w którym po kilku sesjach treningowych bodźce zmieniały się tak, że poprzedni sygnał go stawał się sygnałem no-go (wymagającym większego hamowania). Stwierdzili oni specyficzną aktywację bocznej części BA 8 podczas tych prób. W podobnym badaniu Albares i wsp. (2014), wykorzystali wersję zadania go/no-go, w której przed bodźcami go i no-go prezentowano prime. Jednakże, podczas gdy zielone światło wskazywało, że nastąpi bodziec go, czerwone światło oznaczało, że może wystąpić jeden z trzech warunków: bodziec go, bodziec no-go lub brak bodźca w ogóle. Badacze rozumowali, że warunek czerwonego światła obejmuje bardziej złożone planowanie motoryczne niż zielone światło, i odkryli, że boczne BA 8 było rzeczywiście bardziej aktywne podczas tego warunku.
ADHD i Motor Selekcja i Hamowanie
Są solidne dowody na wybór motoryczny i deficyty hamowania u dzieci z ADHD. Wiele dotychczasowych badań wykazało, że dzieci, młodzież i dorośli z ADHD wykazują gorsze wyniki w różnych wersjach zadania sygnału stop (np. Rubia i in., 1998; Lee i in., 2016; Białostocki i in., 2017; Dekkers i in., 2017), oraz że gdy dzieci z ADHD są odpowiednio leczone i motywowane za pomocą skutecznych wzmocnień, różnice te mogą zanikać (Rosch i in., 2016). Ponadto w bardzo dużym badaniu przeprowadzonym przez Crosbie i wsp. (2013) zmierzono objawy ADHD u ponad 16 000 dzieci ogólnodostępnych, które wykonały SST. Wyniki wykazały, że osoby z większymi wynikami cech ADHD miały gorsze hamowanie reakcji, wolniejszą latencję reakcji i większą zmienność, a także, że tendencja ta była wysoce dziedziczna. Podobnie Alderson et al. (2007), przeprowadzili metaanalizę skupiając się na dzieciach z ADHD i SST i znaleźli większe średnie czasy odpowiedzi, większą zmienność odpowiedzi i większy czas reakcji sygnału stopu w grupie ADHD.
ADHD, Motor Selection, i Inhibition i Lateral BA 8
Istnieją również znaczące dowody wskazujące na specyficzne atypowości w bocznej aktywacji BA 8 w ADHD uczestników wykonujących zadania, które wymagają wyboru silnika i hamowania. Na przykład, dwie oddzielne metaanalizy przeanalizowały badania fMRI oceniające hamowanie motoryczne u dzieci i dorosłych z ADHD (Hart i in., 2013; Rubia i in., 2013). W obu metaanalizach stwierdzono, że niższa aktywacja w bocznym BA 8 jest związana z pomiarami hamowania motorycznego, a w badaniu Rubii i wsp. (2013), stwierdzono również, że metylofenidat zwiększa aktywację w bocznym BA 8, co było związane z lepszym wykonaniem zadań motorycznych. Ponadto w badaniach, w których stosowano różne wersje SST i zadania go/no-go, stwierdzono mniejszą aktywację w bocznym BA 8 w grupie ADHD podczas prostego warunku no-go (Passarotti i in., 2010b; Mulligan i in., 2011), na chwilę przed wystąpieniem błędu (Spinelli i in., 2011), a także podczas warunku wolnego wyboru, w którym uczestnicy wybierali, czy nacisnąć przycisk, czy nie, gdy prezentowany był bodziec (Karch i in., 2010).
Podsumowanie
Bieżąca praca przedstawia dowody na to, że boczna część kory przedczołowej jest zaangażowana w hamowanie, selekcję i manipulację informacji motorycznych, emocjonalnych, pamięciowych i percepcyjno-zmysłowych, funkcję, która jest pomocna w codziennym życiu poprzez zdolność do ignorowania rozproszenia uwagi, wybierania i pobierania określonych informacji z naszej pamięci, identyfikowania i regulowania naszego stanu emocjonalnego lub planowania adekwatnej do sytuacji reakcji motorycznej. Wykorzystuje ten model do klasyfikacji wyzwań napotykanych przez osoby z ADHD, łącząc różne rodzaje trudności w selekcji/hamowaniu z różnymi podobszarami bocznej kory przedczołowej.
Wkład autora
Autor potwierdza, że jest jedynym współautorem tej pracy i zatwierdził ją do publikacji.
Conflict of Interest Statement
Autor oświadcza, że badania zostały przeprowadzone przy braku jakichkolwiek komercyjnych lub finansowych relacji, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Albares, M., Lio, G., Criaud, M., Anton, J. L., Desmurget, M., and Boulinguez, P. (2014). The dorsal medial frontal cortex mediates automatic motor inhibition in uncertain contexts: evidence from combined fMRI and EEG studies. Hum. Brain Mapp. 35, 5517-5531. doi: 10.1002/hbm.22567
PubMed Abstract | CrossRef Full Text | Google Scholar
Alderson, R. M., Rapport, M. D., and Kofler, M. J. (2007). Attention-deficit/hyperactivity disorder and behavioral inhibition: a meta-analytic review of the stop-signal paradigm. J. Abnorm. Child Psychol. 35, 745-758. doi: 10.1007/s10802-007-9131-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Barkley, R. A. (1997). Hamowanie behawioralne, podtrzymywanie uwagi i funkcje wykonawcze: konstruowanie jednolitej teorii ADHD. Psychol. Bull. 121:65. doi: 10.1037/0033-2909.121.1.65
PubMed Abstract | CrossRef Full Text | Google Scholar
Beckmann, M., Johansen-Berg, H., and Rushworth, M. F. (2009). Connectivity-based parcellation of human cingulate cortex and its relation to functional specialization. J. Neurosci. 29, 1175-1190. doi: 10.1523/JNEUROSCI.3328-08.2009
PubMed Abstract | CrossRef Full Text | Google Scholar
Bialystok, E., Hawrylewicz, K., Wiseheart, M., and Toplak, M. (2017). Interakcja dwujęzyczności i zespołu nadpobudliwości psychoruchowej z deficytem uwagi (attention-deficit/hyperactivity disorder) u młodych dorosłych. Bilingualism 20, 588-601. doi: 10.1017/S1366728915000887
PubMed Abstract | CrossRef Full Text | Google Scholar
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., et al. (2014). Cognitive reappraisal of emotion: a meta-analysis of human neuroimaging studies. Cereb. Cortex 24, 2981-2990. doi: 10.1093/cercor/bht154
PubMed Abstract | CrossRef Full Text | Google Scholar
Castel, A. D., Lee, S. S., Humphreys, K. L., and Moore, A. N. (2011). Memory capacity, selective control, and value-directed remembering in children with and without attention-deficit/hyperactivity disorder (ADHD). Neuropsychology 25:15. doi: 10.1037/a0020298
PubMed Abstract | CrossRef Full Text | Google Scholar
Cohen, M. S., Rissman, J., Suthana, N. A., Castel, A. D., and Knowlton, B. J. (2014). Value-based modulation of memory encoding involves strategic engagement of fronto-temporal semantic processing regions. Cogn. Affect. Behav. Neurosci. 14, 578-592. doi: 10.3758/s13415-014-0275-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crosbie, J., Arnold, P., Paterson, A., Swanson, J., Dupuis, A., Li, X., et al. (2013). Response inhibition and ADHD traits: correlates and heritability in a community sample. J. Abnorm. Child Psychol. 41, 497-507. doi: 10.1007/s10802-012-9693-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Dekkers, T. J., van Rentergem, J. A. A., Koole, A., van den Wildenberg, W. P., Popma, A., Bexkens, A., et al. (2017). Time-on-task effects in children with and without ADHD: depletion of executive resources or depletion of motivation? Eur. Child Adolesc. Psychiatry. 26, 1-11. doi: 10.1007/s00787-017-1006-y
CrossRef Full Text | Google Scholar
Depue, B. E., Burgess, G. C., Willcutt, E. G., Ruzic, L., and Banich, M. T. (2010). Inhibitory control of memory retrieval and motor processing associated with the right lateral prefrontal cortex: evidence from deficits in individuals with ADHD. Neuropsychologia. 48, 3909-3917. doi: 10.1016/j.neuropsychologia.2010.09.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Enriquez-Geppert, S., Eichele, T., Specht, K., Kugel, H., Pantev, C., and Huster, R. J. (2013). Functional parcellation of the inferior frontal and midcingulate cortices in a flanker-stop-change paradigm. Hum. Brain Mapp. 34, 1501-1514. doi: 10.1002/hbm.22002
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., and Lavie, N. (2016). Ustalenie cechy rozproszenia uwagi (attention-distractibility trait). Psychol. Sci. 27, 203-212. doi: 10.1177/0956797615617761
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., Robertson, D. J., Jennings, A., Asherson, P., and Lavie, N. (2014). Plugging deficytu uwagi: perceptual load counters increased distraction in ADHD. Neuropsychology 28:91. doi: 10.1037/neu0000020
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank, D. W., Dewitt, M., Hudgens-Haney, M., Schaeffer, D. J., Ball, B. H., Schwarz, N. F., et al. (2014). Emotion regulation: quantitative meta-analysis of functional activation and deactivation. Neurosci. Biobehav. Rev. 45, 202-211. doi: 10.1016/j.neubiorev.2014.06.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Gilbert, S. J., Spengler, S., Simons, J. S., Steele, J. D., Lawrie, S. M., Frith, C. D., et al. (2006). Functional specialization within rostral prefrontal cortex (area 10): a meta-analysis. J. Cogn. Neurosci. 18, 932-948. doi: 10.1162/jocn.2006.18.6.932
PubMed Abstract | CrossRef Full Text | Google Scholar
Graziano, P. A., and Garcia, A. (2016). Attention-deficit hyperactivity disorder and children’s emotion dysregulation: a meta-analysis. Clin. Psychol. Rev. 46, 106-123. doi: 10.1016/j.cpr.2016.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Hallam, G. P., Webb, T. L., Sheeran, P., Miles, E., Niven, K., Wilkinson, I. D., et al. (2014). The neural correlates of regulating another person’s emotions: an exploratory fMRI study. Front. Hum. Neurosci. 8:376. doi: 10.3389/fnhum.2014.00376
PubMed Abstract | CrossRef Full Text | Google Scholar
Hart, H., Radua, J., Nakao, T., Mataix-Cols, D., and Rubia, K. (2013). Meta-analysis of functional magnetic resonance imaging studies of inhibition and attention in attention-deficit/hyperactivity disorder: exploring task-specific, stimulant medication, and age effects. JAMA Psychiatry. 70, 185-198. doi: 10.1001/jamapsychiatry.2013.277
PubMed Abstract | CrossRef Full Text | Google Scholar
Howard, J. D., and Kahnt, T. (2017). Identity-specific reward representations in orbitofrontal cortex are modulated by selective devaluation. J. Neurosci. 37, 3473-3416. doi: 10.1523/JNEUROSCI.3473-16.2017
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, M. E., Budd, T. W., Fulham, W. R., Lancaster, S., Woods, W., Rossell, S. L., et al. (2014). Sustained brain activation supporting stop-signal task performance. Eur. J. Neurosci. 39, 1363-1369. doi: 10.1111/ejn.12497
PubMed Abstract | CrossRef Full Text | Google Scholar
Hwang, S., White, S. F., Nolan, Z. T., Craig Williams, W., Sinclair, S., and Blair, R. J. (2015). Executive attention control and emotional responding in attention-deficit/hyperactivity disorder-A functional MRI study. Neuroimage Clin. 9, 545-554. doi: 10.1016/j.nicl.2015.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Ikeda, Y., Okuzumi, H., and Kokubun, M. (2013). Stroop/reverse-Stroop interference in typical development and its relation to symptoms of ADHD. Res. Dev. Disabil. 34, 2391-2398. doi: 10.1016/j.ridd.2013.04.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Karch, S., Thalmeier, T., Lutz, J., Cerovecki, A., Opgen-Rhein, M., Hock, B., et al. (2010). Neural correlates (ERP/fMRI) of voluntary selection in adult ADHD patients. Eur. Arch. Psychiatry Clin. Neurosci. 260, 427-440. doi: 10.1007/s00406-009-0089-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaufman, D. A., Keith, C. M., and Perlstein, W. M. (2016). Orbitofrontal cortex and the early processing of visual novelty in healthy aging. Front. Aging Neurosci. 8:101. doi: 10.3389/fnagi.2016.00101
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2011). Aktywność neuronalna, która przewiduje późniejsze zapamiętywanie i zapominanie: metaanaliza 74 badań fMRI. Neuroimage 54, 2446-2461. doi: 10.1016/j.neuroimage.2010.09.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2013). Zróżnicowana aktywność neuronalna w rozpoznawaniu starych i nowych wydarzeń: metaanaliza estymacji prawdopodobieństwa aktywacji. Hum. Brain Mapp. 34, 814-836. doi: 10.1002/hbm.21474
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. Y., Wu, T. F., Tsai, J. D., and Yang, E. L. (2016). Applicability of the stop-signal task for preschoolers with ADHD. Percept. Mot. Skills. 123, 162-174. doi: 10.1177/0031512516660715
PubMed Abstract | CrossRef Full Text | Google Scholar
Lenartowicz, A., Verbruggen, F., Logan, G. D., and Poldrack, R. A. (2011). Inhibition-related activation in the right inferior frontal gyrus in the absence of inhibitory cues. J. Cogn. Neurosci. 23, 3388-3399. doi: 10.1162/jocn_a_00031
PubMed Abstract | CrossRef Full Text | Google Scholar
Matthews, S. C., Simmons, A. N., Arce, E., and Paulus, M. P. (2005). Dissociation of inhibition from error processing using a parametric inhibitory task during functional magnetic resonance imaging. Neuroreport. 16, 755-760. doi: 10.1097/00001756-200505120-00020
PubMed Abstract | CrossRef Full Text | Google Scholar
Mulligan, R. C., Knopik, V. S., Sweet, L. H., Fischer, M., Seidenberg, M., and Rao, S. M. (2011). Neural correlates of inhibitory control in adult attention deficit/hyperactivity disorder: evidence from the Milwaukee longitudinal sample. Psychiatry Res. 194, 119-129. doi: 10.1016/j.pscychresns.2011.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Nigg, J. T. (1999). Deficyt zahamowania reakcji ADHD mierzony przez zadanie zatrzymania: replikacja z DSM-IV połączonego typu, rozszerzenia i kwalifikacji. J. Abnorm. Child Psychol. 27, 393-402. doi: 10.1023/A:1021980002473
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010a). Neural correlates of response inhibition in pediatric bipolar disorder and attention deficit hyperactivity disorder. Psychiatry Res. Neuroimaging. 181, 36-43. doi: 10.1016/j.pscychresns.2009.07.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010b). Differential engagement of cognitive and affective neural systems in pediatric bipolar disorder and attention deficit hyperactivity disorder. J. Int. Neuropsychol. Soc. 16, 106-117. doi: 10.1017/S1355617709991019
PubMed Abstract | CrossRef Full Text | Google Scholar
Pelletier, M. F., Hodgetts, H. M., Lafleur, M. F., Vincent, A., and Tremblay, S. (2016). Podatność na efekt nieistotnego dźwięku u dorosłych z ADHD. J. Atten. Disord. 20, 306-316. doi: 10.1177/1087054713492563
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollak, Y., Kahana-Vax, G., and Hoofien, D. (2007). Procesy wyszukiwania informacji u dorosłych z ADHD: badanie RAVLT. Dev. Neuropsychol. 33, 62-73. doi: 10.1080/87565640701729789
PubMed Abstract | CrossRef Full Text | Google Scholar
Pretus, C., Picado, M., Ramos-Quiroga, A., Carmona, S., Richarte, V., Fauquet, J., et al. (2016). Presence of distractor improves time estimation performance in an adult ADHD Sample. J. Atten. Disord. doi: 10.1177/1087054716648776. .
PubMed Abstract | CrossRef Full Text | Google Scholar
Price, J. L. (2007). Definicja kory oczodołowej w odniesieniu do specyficznych połączeń ze strukturami limbicznymi i trzewnymi oraz innymi regionami korowymi. Annals N.Y. Acad. Sci. 1121, 54-71. doi: 10.1196/annals.1401.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabinak, C. A., MacNamara, A., Kennedy, A. E., Angstadt, M., Stein, M. B., Liberzon, I., et al. (2014). Focal and aberrant prefrontal engagement during emotion regulation in veterans with posttraumatic stress disorder. Depress. Anxiety. 31, 851-861. doi: 10.1002/da.22243
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, B., Mehler, B., D’Ambrosio, L. A., and Fried, R. (2010). The impact of distractions on young adult drivers with attention deficit hyperactivity disorder (ADHD). Accid. Anal. Prev. 42, 842-851. doi: 10.1016/j.aap.2009.06.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosch, K. S., Fosco, W. D., Pelham, W. E. Jr, Waxmonsky, J. G., Bubnik, M. G., and Hawk, L. W. Jr (2016). Reinforcement and stimulant medication ameliorate deficient response inhibition in children with attention-deficit/hyperactivity disorder. J. Abnorm. Child Psychol. 44, 309-321. doi: 10.1007/s10802-015-0031-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Lim, L., Ecker, C., Halari, R., Giampietro, V., Simmons, A., et al. (2013). Effects of age and gender on neural networks of motor response inhibition: from adolescence to mid-adulthood. Neuroimage. 83, 690-703. doi: 10.1016/j.neuroimage.2013.06.078
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Oosterlaan, J., Sergeant, J. A., Brandeis, D., and Leeuwen, T. V. (1998). Inhibitory dysfunction in hyperactive boys. Behav. Brain Res. 94, 25-32. doi: 10.1016/S0166-4328(97)00166-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Shalom, D. B. (2009). Przyśrodkowa kora przedczołowa i integracja w autyzmie. Neuroscientist. 15, 589-598. doi: 10.1177/1073858409336371
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, J. L., Jamadar, S., Provost, A. L., and Michie, P. T. (2013). Motoryczne i nie-motoryczne hamowanie w zadaniu Go/NoGo: badanie ERP i fMRI. Int. J. Psychophysiol. 87, 244-253. doi: 10.1016/j.ijpsycho.2012.07.185
PubMed Abstract | CrossRef Full Text | Google Scholar
Soliman, A. M., and Elfar, R. M. (2017). Fałszywa pamięć u dorosłych z ADHD: porównanie między podtypami i normalnymi kontrolami. J. Attent. Disord. 21, 986-996. doi: 10.1177/1087054714556814
PubMed Abstract | CrossRef Full Text | Google Scholar
Sørensen, L., Plessen, K. J., Adolfsdottir, S., and Lundervold, A. J. (2014). The specificity of the Stroop interference score of errors to ADHD in boys. Child Neuropsychol. 20, 677-691. doi: 10.1080/09297049.2013.855716
PubMed Abstract | CrossRef Full Text | Google Scholar
Soto, D., Humphreys, G. W., and Rotshtein, P. (2007). Dissociating the neural mechanisms of memory-based guidance of visual selection. Proc. Natl. Acad. Sci. U.S.A. 104, 17186-17191. doi: 10.1073/pnas.0703706104
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinelli, S., Joel, S., Nelson, T. E., Vasa, R. A., Pekar, J. J., and Mostofsky, S. H. (2011). Different neural patterns are associated with trials preceding inhibitory errors in children with and without attention-deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry. 50, 705-715. doi: 10.1016/j.jaac.2011.03.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsujimoto, S., Yasumura, A., Yamashita, Y., Torii, M., Kaga, M., and Inagaki, M. (2013). Increased prefrontal oxygenation related to distractor-resistant working memory in children with attention-deficit/hyperactivity disorder (ADHD). Child Psychiatry Hum. Dev. 44, 678-688. doi: 10.1007/s10578-013-0361-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcott, C. M., and Landau, S. (2004). The relationship between disinhibition and emotion regulation in boys with attention deficit hyperactivity disorder. J. Clin. Child Adolesc. Psychol. 33, 772-782. doi: 10.1207/s15374424jccp3304_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Xiong, K., Zhang, Y., Qiu, M., Zhang, J., Sang, L., Wang, L., et al. (2013). Negative emotion regulation in patients with posttraumatic stress disorder. PLoS ONE 8:e81957. doi: 10.1371/journal.pone.0081957
PubMed Abstract | CrossRef Full Text | Google Scholar
Yarmolovsky, J., Szwarc, T., Schwartz, M., Tirosh, E., and Geva, R. (2016). Gorąca kontrola wykonawcza i odpowiedź na stymulant w podwójnie ślepym randomizowanym badaniu u dzieci z ADHD. Eur. Arch. Psychiatry Clin. Neurosci. 267, 73-82. doi: 10.1007/s00406-016-0683-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Yasumura, A., Kokubo, N., Yamamoto, H., Yasumura, Y., Nakagawa, E., Kaga, M., et al. (2014). Neurobehawioralna i hemodynamiczna ocena interferencji Stroopa i odwróconego Stroopa u dzieci z zaburzeniami koncentracji uwagi/hiperaktywności. Brain Dev. 36, 97-106. doi: 10.1016/j.braindev.2013.01.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Zsuga, J., Biro, K., Tajti, G., Szilasi, M. E., Papp, C., Juhasz, B., et al. (2016). 'Proactive’ use of cue-context congruence for building reinforcement learning’s reward function. BMC Neurosci. 17:70. doi: 10.1186/s12868-016-0302-7
PubMed Abstract | CrossRef Full Text | Google Scholar
.