Eukariota ma dwa główne typy podziału komórek: mitozę i mejozę. Mitoza jest używana do produkcji nowych komórek ciała do wzrostu i leczenia, podczas gdy mejoza jest używana do produkcji komórek płciowych (jaj i plemników). Mejoza zostanie omówiona w późniejszym rozdziale.
Cykl komórkowy jest uporządkowaną serią zdarzeń obejmujących wzrost i podział komórki, która produkuje dwie nowe komórki córki poprzez mitozę. Długość cyklu komórkowego jest bardzo zmienna, nawet w obrębie komórek pojedynczego organizmu. U ludzi częstotliwość obrotu komórkowego waha się od kilku godzin we wczesnym rozwoju embrionalnym do średnio dwóch do pięciu dni w przypadku komórek nabłonkowych, lub do całego życia ludzkiego spędzonego bez podziału w wyspecjalizowanych komórkach, takich jak neurony korowe lub komórki mięśnia sercowego. Różny jest również czas, jaki komórka spędza w każdej fazie cyklu komórkowego. Kiedy szybko dzielące się komórki ssaków są hodowane w kulturach (poza organizmem w optymalnych warunkach wzrostu), długość cyklu wynosi około 24 godzin. Czas wydarzeń w cyklu komórkowym jest kontrolowany przez mechanizmy, które są zarówno wewnętrzne, jak i zewnętrzne w stosunku do komórki.
Komórki na drodze do podziału komórkowego przechodzą przez serię precyzyjnie określonych w czasie i starannie regulowanych etapów wzrostu, replikacji DNA i podziału, które produkują dwie genetycznie identyczne komórki. Cykl komórkowy ma dwie główne fazy: interfazę i fazę mitotyczną (rysunek 1). Podczas fazy międzyfazowej komórka rośnie i replikuje DNA. Podczas fazy mitotycznej replikowane DNA i zawartość cytoplazmy są rozdzielane, a komórka dzieli się.
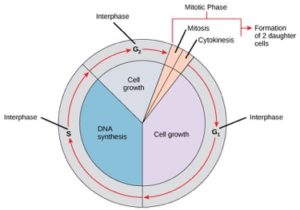
Podczas interfazy komórka przechodzi normalne procesy, przygotowując się jednocześnie do podziału komórki. Aby komórka mogła przejść z fazy interfazowej do mitotycznej, musi być spełnionych wiele warunków wewnętrznych i zewnętrznych. Trzy etapy interfazy są nazywane G1, S i G2.
Faza G1 (pierwsza luka)
Pierwszy etap interfazy jest nazywany fazą G1 (pierwsza luka), ponieważ z mikroskopowego punktu widzenia niewiele zmian jest widocznych. Jednakże, podczas etapu G1, komórka jest dość aktywny na poziomie biochemicznym. Komórka gromadzi składniki budulcowe chromosomalnego DNA i związanych z nim białek, jak również gromadzi wystarczające rezerwy energii, aby wykonać zadanie replikacji każdego chromosomu w jądrze.
Faza S (synteza DNA)
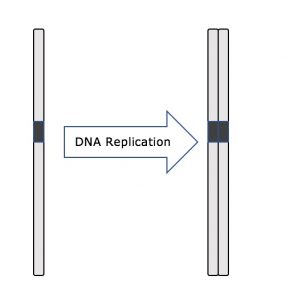
Przez całą interfazę, jądrowe DNA pozostaje w konfiguracji półzwartej chromatyny. W fazie S replikacja DNA może przebiegać poprzez mechanizmy, które prowadzą do powstania identycznych par cząsteczek DNA – siostrzanych chromatyd – które są mocno przytwierdzone do regionu centromerowego (Rysunek 2).
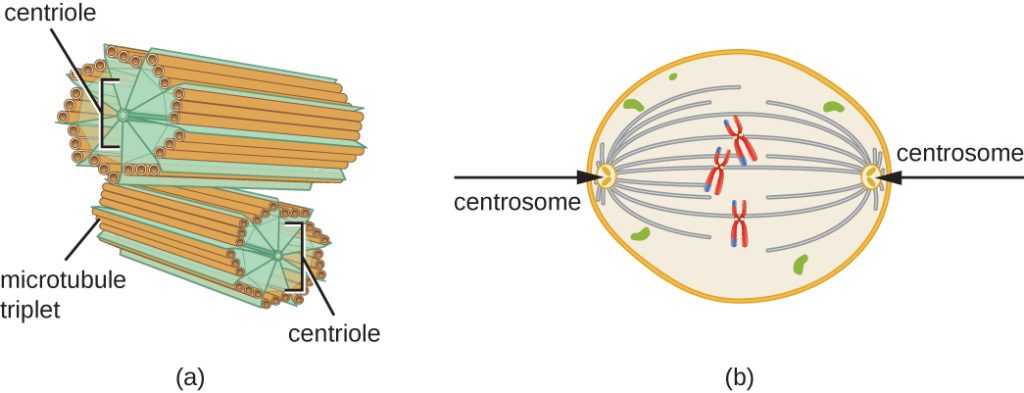
W fazie S powielany jest również centrosom. Te dwa centrosomy dadzą początek wrzecionu mitotycznemu, aparatowi, który organizuje ruch chromosomów podczas mitozy. W centrum każdej komórki zwierzęcej, centrosomy komórek zwierzęcych są związane z parą pręcikowatych obiektów, centrioli, które są ustawione względem siebie pod kątem prostym. Centriole pomagają organizować podziały komórkowe. Centriole nie są obecne w centrosomach innych gatunków eukariotycznych, takich jak rośliny i większość grzybów.
Faza G2 (Second Gap)
W fazie G2 komórka uzupełnia swoje zapasy energii i syntetyzuje białka niezbędne do manipulacji chromosomami. Niektóre organelle komórkowe są duplikowane, a cytoszkielet jest demontowany w celu zapewnienia zasobów dla fazy mitotycznej. Podczas G2 może nastąpić dodatkowy wzrost komórek. Ostateczne przygotowania do fazy mitotycznej muszą być zakończone, zanim komórka będzie mogła wejść w pierwszy etap mitozy.
Faza mitotyczna
Aby powstały dwie komórki córki, zawartość jądra i cytoplazmy musi zostać podzielona. Faza mitotyczna jest wieloetapowym procesem, podczas którego zduplikowane chromosomy są wyrównywane, rozdzielane i przemieszczane do przeciwległych biegunów komórki, a następnie komórka dzieli się na dwie nowe, identyczne komórki córki. Pierwsza część fazy mitotycznej, mitoza, składa się z pięciu etapów, które realizują podział jądrowy (rysunek 5). Druga część fazy mitotycznej, zwana cytokinezą, polega na fizycznym rozdzieleniu składników cytoplazmatycznych na dwie komórki potomne. Chociaż etapy mitozy są podobne u większości eukariotów, proces cytokinezy jest zupełnie inny u eukariotów posiadających ściany komórkowe, takich jak komórki roślinne.
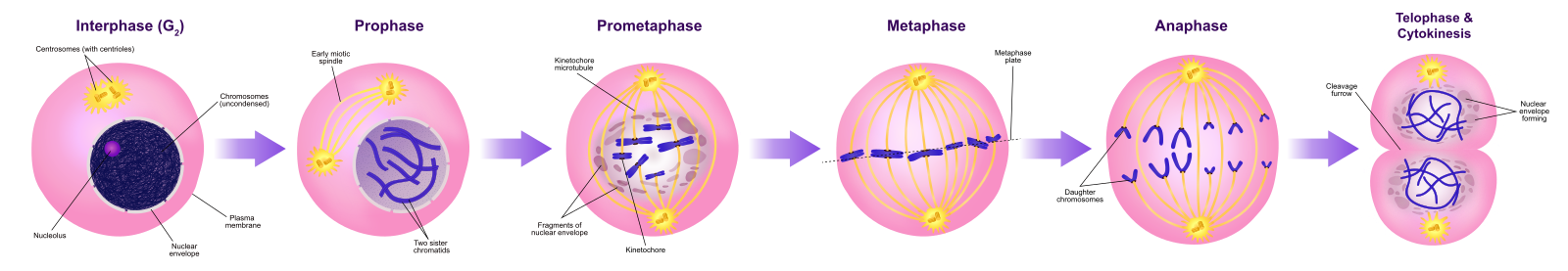
Profazy
Podczas profazy, „pierwszej fazy”, otoczka jądrowa zaczyna rozpadać się na małe pęcherzyki, a organelle błoniaste (takie jak aparat Golgiego i retikulum endoplazmatyczne), fragmentują się i rozpraszają w kierunku brzegów komórki. Zanika jąderko. Centrosomy zaczynają przemieszczać się do przeciwległych biegunów komórki. Mikrotubule, które utworzą wrzeciono mitotyczne, rozciągają się pomiędzy centrosomami, przesuwając je dalej od siebie w miarę wydłużania się włókien mikrotubul. Chromatydy siostrzane zaczynają się ciaśniej zwijać przy pomocy białek kondensynowych i stają się widoczne pod mikroskopem świetlnym.
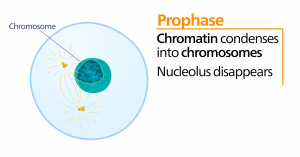
Prometafaza
Podczas prometafazy, „pierwszej fazy przemiany”, wiele procesów, które zostały rozpoczęte w profazie, nadal postępuje. Pozostałości otoczki jądrowej ulegają fragmentacji. Wrzeciono mitotyczne nadal się rozwija, ponieważ więcej mikrotubul gromadzi się i rozciąga na całej długości dawnego obszaru jądrowego. Chromosomy stają się bardziej skondensowane i dyskretne. Każda chromatyda siostrzana rozwija strukturę białkową zwaną kinetochorem w regionie centromerowym.
Białka kinetochoru przyciągają i wiążą mikrotubule wrzeciona mitotycznego. Gdy mikrotubule wrzeciona wysuwają się z centrosomów, niektóre z tych mikrotubul stykają się z kinetochorami i mocno się z nimi wiążą. Gdy włókno mitotyczne przyczepi się do chromosomu, chromosom ten będzie zorientowany tak, że kinetochory chromatyd siostrzanych będą zwrócone ku przeciwległym biegunom. Ostatecznie wszystkie chromatydy siostrzane zostaną przyłączone poprzez swoje kinetochore do mikrotubul z przeciwległych biegunów. Mikrotubule wrzecionowe, które nie przylegają do chromosomów nazywane są mikrotubulami polarnymi. Mikrotubule te zachodz± na siebie w połowie drogi między dwoma biegunami i przyczyniaj± się do wydłużania komórki. Mikrotubule astralne znajdują się w pobliżu biegunów, pomagają w orientacji wrzeciona i są wymagane do regulacji mitozy.

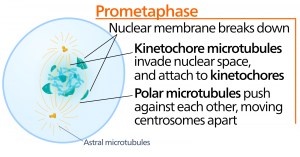
Metafaza
Podczas metafazy, „fazy przemiany”, wszystkie chromosomy są ułożone w płaszczyźnie zwanej płytką metafazową lub płaszczyzną równikową, w połowie drogi między dwoma biegunami komórki. Chromatydy siostrzane są nadal ściśle połączone ze sobą przez białka kohezyny. W tym czasie chromosomy są maksymalnie skondensowane.
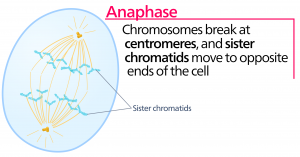
Anafaza
Podczas anafazy, „fazy wznoszącej”, białka kohezynowe ulegają degradacji, a chromatydy siostrzane rozdzielają się w centromerze. Każda chromatyda, zwana teraz chromosomem, jest gwałtownie ciągnięta w kierunku centrosomu, do którego przyczepiona jest jej mikrotubula. Komórka staje się widocznie wydłużona (owalna), ponieważ mikrotubule biegunowe ślizgają się względem siebie na płytce metafazy, gdzie zachodzą na siebie.
Telofaza
Podczas telofazy, „fazy dystansowej”, chromosomy osiągają przeciwległe bieguny i zaczynają się dekondensować (rozpraszać), rozluźniając się do konfiguracji chromatyny. Wrzeciona mitotyczne ulegają depolimeryzacji do monomerów tubuliny, które zostaną wykorzystane do złożenia elementów cytoszkieletu dla każdej komórki potomnej. Otoczki jądrowe tworzą się wokół chromosomów, a nukleosomy pojawiają się w obrębie jądra.
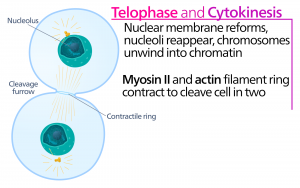
Cytokineza
Cytokineza, czyli „ruch komórki”, jest drugim głównym etapem fazy mitotycznej, podczas którego podział komórki zostaje zakończony poprzez fizyczne rozdzielenie składników cytoplazmatycznych na dwie komórki potomne. Podział nie jest zakończony, dopóki składniki komórki nie zostaną podzielone i całkowicie rozdzielone na dwie komórki córki. Chociaż etapy mitozy są podobne dla większości eukariotów, proces cytokinezy jest zupełnie inny dla eukariotów, które mają ściany komórkowe, takie jak komórki roślinne.
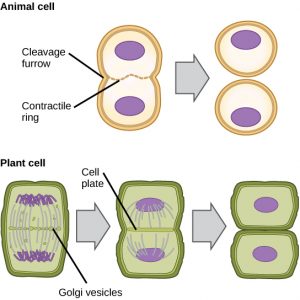
W komórkach takich jak komórki zwierzęce, które nie mają ścian komórkowych, cytokineza następuje po rozpoczęciu anafazy. Kurczliwy pierścień złożony z włókien aktyny tworzy się tuż wewnątrz błony plazmatycznej przy dawnej płytce metafazowej (Rysunek 12). Włókna aktyny wciągają równik komórki do środka, tworząc szczelinę. Ta szczelina, czyli „pęknięcie”, nazywana jest bruzdą cięcia. Bruzda pogłębia się w miarę kurczenia się pierścienia aktynowego i ostatecznie błona zostaje rozcięta na dwie części.
W komórkach roślinnych nowa ściana komórkowa musi powstać pomiędzy komórkami córkami. Podczas interfazy, aparat Golgiego gromadzi enzymy, białka strukturalne i cząsteczki glukozy przed rozbiciem na pęcherzyki i rozproszeniem w całej dzielącej się komórce (Rysunek 12). Podczas telofazy te pęcherzyki Golgiego są transportowane na mikrotubulach w celu utworzenia fragmoplastu (struktury pęcherzykowej) na płytce metafazowej. Tam pęcherzyki łączą się i koalescendują od środka w kierunku ścian komórkowych; struktura ta nazywana jest płytką komórkową. W miarę jak łączą się kolejne pęcherzyki, płytka komórkowa powiększa się, aż połączy się ze ścianami komórkowymi na obrzeżach komórki. Enzymy wykorzystują glukozę, która nagromadziła się pomiędzy warstwami błon, do budowy nowej ściany komórkowej. Błony Golgiego stają się częściami błony plazmatycznej po obu stronach nowej ściany komórkowej.
Podsumowanie Mitozy i Cytokinezy
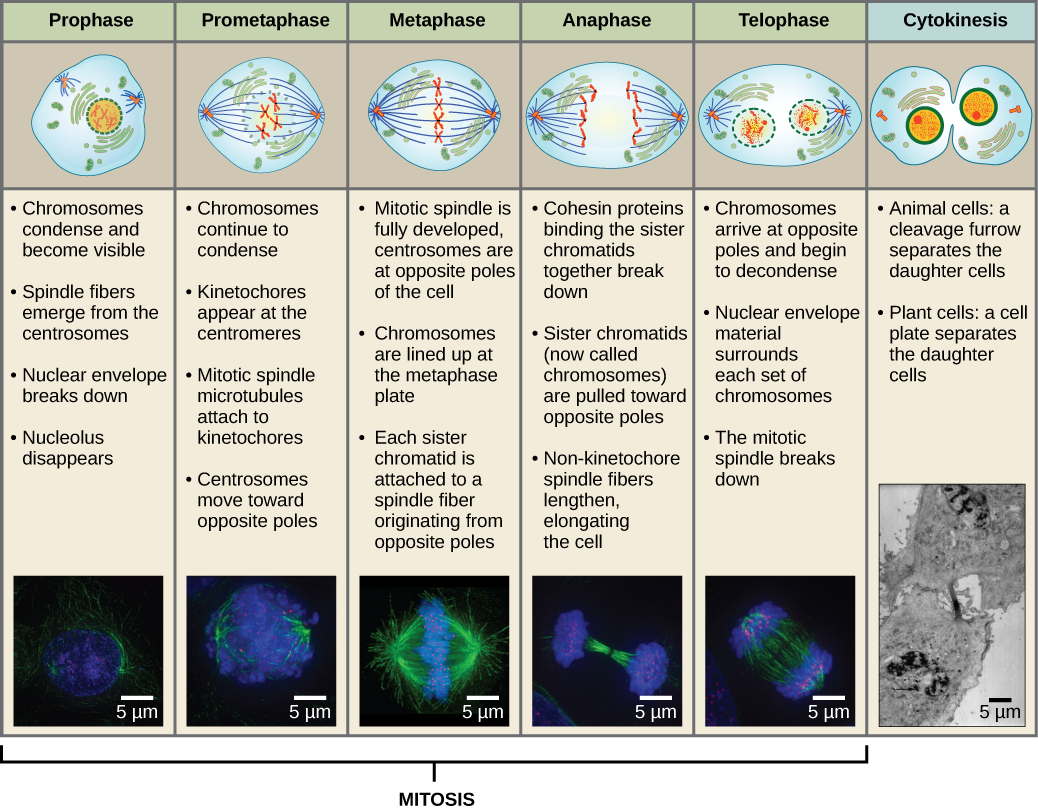
Rysunek 13 Mitoza dzieli się na pięć etapów-profazę, prometafazę, metafazę, anafazę i telofazę. Zdjęcia u dołu zostały wykonane za pomocą mikroskopii fluorescencyjnej komórek sztucznie wybarwionych barwnikami fluorescencyjnymi: niebieska fluorescencja oznacza DNA (chromosomy), a zielona fluorescencja mikrotubule (aparat wrzecionowy). (kredyt „rysunki mitozy”: modyfikacja pracy Mariany Ruiz Villareal; kredyt „mikrografy”: modyfikacja pracy Roya van Heesbeena; kredyt „mikrograf cytokinezy”: Wadsworth Center/New York State Department of Health; scale-bar data from Matt Russell)
Faza G0
Nie wszystkie komórki przylegają do klasycznego wzorca cyklu komórkowego, w którym nowo utworzona komórka-córka natychmiast wchodzi w interfazę, po której następuje faza mitotyczna. Komórki w fazie G0 nie są aktywnie przygotowywane do podziału. Komórka jest w fazie quiescent (nieaktywnej), po wyjściu z cyklu komórkowego. Niektóre komórki wchodzą w fazę G0 tymczasowo, dopóki zewnętrzny sygnał nie spowoduje rozpoczęcia fazy G1. Inne komórki, które nigdy lub rzadko się dzielą, takie jak dojrzały mięsień sercowy i komórki nerwowe, pozostają w G0 na stałe).