Eukaryoter har två huvudtyper av celldelning: mitos och meios. Mitos används för att producera nya kroppsceller för tillväxt och läkning, medan meios används för att producera könsceller (ägg och spermier). Meiosen kommer att diskuteras i ett senare kapitel.
Cellcykeln är en ordnad serie händelser som involverar celltillväxt och celldelning som producerar två nya dotterceller via mitos. Cellcykelns längd är mycket varierande även inom cellerna i en enskild organism. Hos människor varierar frekvensen av cellomsättning från några timmar under tidig embryonal utveckling till i genomsnitt två till fem dagar för epitelceller, eller till ett helt människoliv som tillbringas utan att dela sig i specialiserade celler som kortikala neuroner eller hjärtmuskelceller. Det finns också en variation i den tid som en cell tillbringar i varje fas av cellcykeln. När snabbdelande däggdjursceller odlas i kultur (utanför kroppen under optimala odlingsförhållanden) är cykelns längd ungefär 24 timmar. Tidpunkten för händelserna i cellcykeln styrs av mekanismer som är både interna och externa till cellen.
Celler på väg mot celldelning går igenom en serie exakt tidsbestämda och noggrant reglerade stadier av tillväxt, DNA-replikation och delning som ger två genetiskt identiska celler. Cellcykeln har två huvudfaser: interfasen och mitosfasen (figur 1). Under interfasen växer cellen och DNA replikeras. Under den mitotiska fasen separeras det replikerade DNA:t och cytoplasmainnehållet och cellen delar sig.
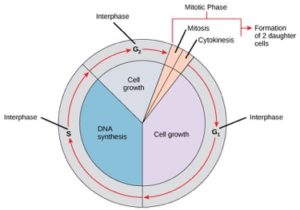
Under interfasen genomgår cellen normala processer samtidigt som den förbereder sig för celldelning. För att en cell ska kunna gå från interfasen till mitosfasen måste många interna och externa villkor vara uppfyllda. De tre stegen i interfasen kallas G1, S och G2.
G1-fasen (första gapet)
Det första steget i interfasen kallas G1-fasen (första gapet) eftersom det ur mikroskopisk synvinkel inte syns någon större förändring. Under G1-stadiet är dock cellen ganska aktiv på biokemisk nivå. Cellen ackumulerar byggstenarna i kromosomalt DNA och de associerade proteinerna samt ackumulerar tillräckliga energireserver för att slutföra uppgiften att replikera varje kromosom i kärnan.
S-fasen (DNA-syntes)
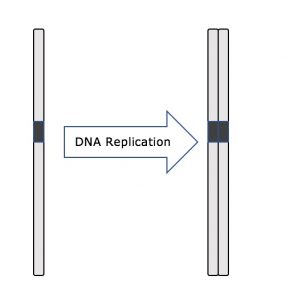
Under hela interfasen förblir kärn-DNA i en halvkondenserad kromatinkonfiguration. I S-fasen kan DNA-replikationen fortsätta genom de mekanismer som resulterar i bildandet av identiska par av DNA-molekyler – systerkromatider – som är fast knutna till den centromeriska regionen (figur 2).
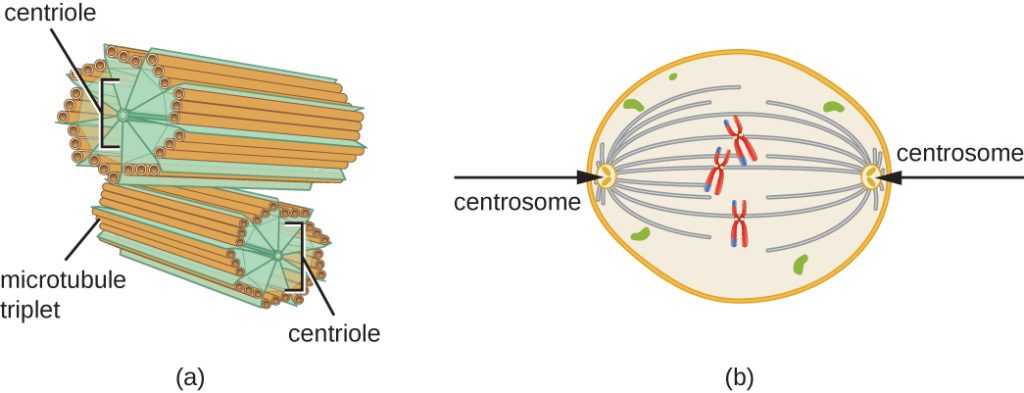
Centrosomen dupliceras också under S-fasen. De två centrosomerna kommer att ge upphov till den mitotiska spindeln, den apparat som iscensätter kromosomernas rörelse under mitosen. I centrum av varje djurcell är djurcellernas centrosomer förknippade med ett par stavliknande objekt, centriolerna, som står i rät vinkel mot varandra. Centriolerna hjälper till att organisera celldelningen. Centrioler finns inte i centrosomerna hos andra eukaryota arter, såsom växter och de flesta svampar.
G2-fas (andra gapet)
I G2-fasen fyller cellen på sina energidepåer och syntetiserar proteiner som är nödvändiga för kromosommanipulation. Vissa cellorganeller dupliceras och cytoskelettet monteras ned för att tillhandahålla resurser för mitosfasen. Det kan förekomma ytterligare celltillväxt under G2. De sista förberedelserna för mitosfasen måste vara avslutade innan cellen kan gå in i mitosens första steg.
Den mitotiska fasen
För att skapa två dotterceller måste innehållet i kärnan och cytoplasman delas. Mitosfasen är en process i flera steg under vilken de duplicerade kromosomerna anpassas, separeras och flyttas till cellens motsatta poler, varefter cellen delas i två nya identiska dotterceller. Den första delen av den mitotiska fasen, mitos, består av fem steg som genomför kärndelningen (figur 5). Den andra delen av mitosfasen, kallad cytokinesis, är den fysiska uppdelningen av cytoplasmakomponenterna i två dotterceller. Även om mitosens faser är likartade för de flesta eukaryoter är cytokinesisprocessen helt annorlunda för eukaryoter som har cellväggar, till exempel växtceller.
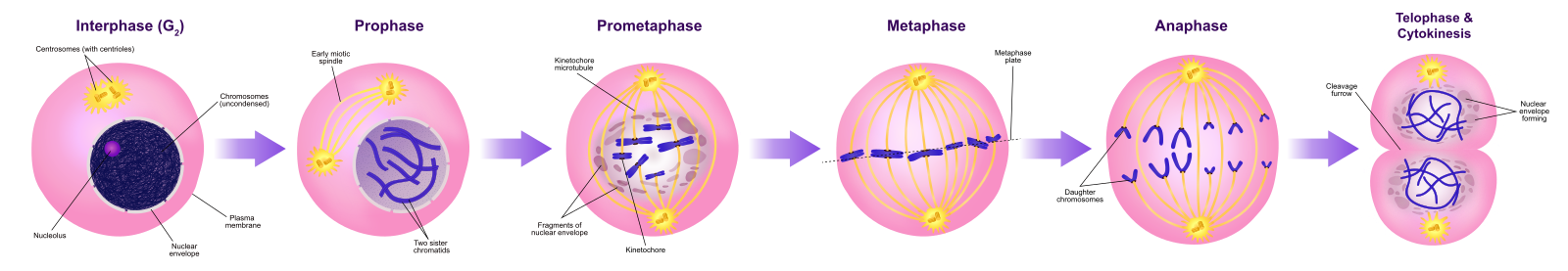
Profas
Under profasen, den ”första fasen”, börjar kärnhöljet att dissociera i små vesiklar, och de membranösa organellerna (såsom Golgiapparaten och det endoplasmatiska retikulumet), fragmenteras och sprids mot cellens kanter. Nukleolus försvinner. Centrosomerna börjar flytta sig till cellens motsatta poler. Mikrotubuli som kommer att bilda den mitotiska spindeln sträcker sig mellan centrosomerna och pressar dem längre ifrån varandra när mikrotubulfibrerna förlängs. Systerkromatiderna börjar rulla ihop sig tätare med hjälp av kondensinproteiner och blir synliga i ett ljusmikroskop.
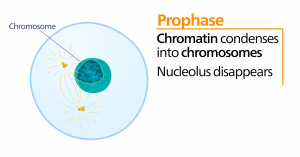
Prometafas
Under prometafas, den ”första förändringsfasen”, fortsätter många processer som påbörjades i profasen att gå framåt. Resterna av kärnhöljet fragmenterar. Den mitotiska spindeln fortsätter att utvecklas när fler mikrotubuli samlas och sträcker sig över längden av det tidigare kärnområdet. Kromosomerna blir mer kondenserade och diskreta. Varje systerkromatid utvecklar en proteinstruktur som kallas kinetokor i den centromeriska regionen.
Kinetokorens proteiner attraherar och binder mitotisk spindelmikrotubuli. När spindelns mikrotubuli sträcker sig från centrosomerna kommer en del av dessa mikrotubuli i kontakt med och binder fast till kinetokorerna. När en mitotisk fiber fäster vid en kromosom kommer kromosomen att orienteras tills systerkromatidernas kinetokorer står mot motsatta poler. Så småningom kommer alla systerkromatider att fästas via sina kinetokorer vid mikrotubuli från motsatta poler. Spindelns mikrotubuli som inte griper in i kromosomerna kallas polära mikrotubuli. Dessa mikrotubuli överlappar varandra halvvägs mellan de två polerna och bidrar till cellförlängningen. Astrala mikrotubuli finns nära polerna, hjälper till med spindelorientering och krävs för reglering av mitos.

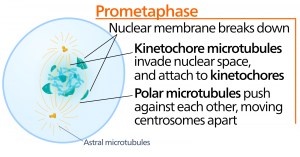
Metafas
Under metafas, ”förändringsfasen”, är alla kromosomer uppradade i ett plan som kallas metafasplattan, eller ekvatorialplanet, mitt emellan cellens två poler. Systerkromatiderna är fortfarande fast knutna till varandra med hjälp av kohesinproteiner. Vid denna tidpunkt är kromosomerna maximalt kondenserade.
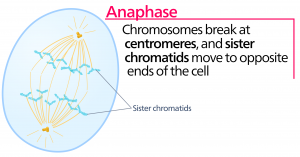
Anafas
Under anafasen, den ”uppåtgående fasen”, bryts kohesinproteinerna ner och systerkromatiderna separeras vid centromeren. Varje kromatid, som nu kallas kromosom, dras snabbt mot centrosomen till vilken dess mikrotubuli är fästade. Cellen blir synligt förlängd (oval) när de polära mikrotubulerna glider mot varandra vid metafasplattan där de överlappar varandra.
Telofas
Under telofasen, ”distansfasen”, når kromosomerna de motsatta polerna och börjar dekondenseras (avvecklas) och slappnar av till en kromatinkonfiguration. De mitotiska spindlarna depolymeriseras till tubulinmonomerer som kommer att användas för att sätta ihop cytoskelettkomponenter för varje dottercell. Kärnhöljen bildas runt kromosomerna och nukleosomer uppträder inom kärnområdet.
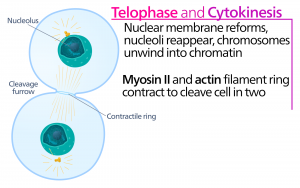
Cytokinesis
Cytokinesis, eller ”cellrörelse”, är det andra huvudstadiet i mitosfasen under vilket celldelningen avslutas via den fysiska separationen av de cytoplasmatiska komponenterna i två dotterceller. Delningen är inte avslutad förrän cellkomponenterna har delats och helt separerats i de två dottercellerna. Även om mitosens stadier är likartade för de flesta eukaryoter är cytokinesiprocessen helt annorlunda för eukaryoter som har cellväggar, t.ex. växtceller.
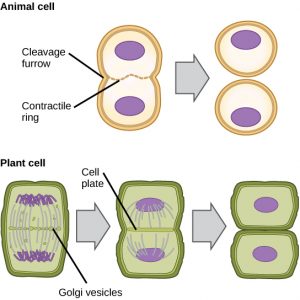
I celler, t.ex. djurceller som saknar cellväggar, följer cytokinesiprocessen efter anafasens början. En kontraktila ring bestående av aktinfilament bildas strax innanför plasmamembranet vid den tidigare metafasplattan (figur 12). Aktinfilamenten drar cellens ekvator inåt och bildar en spricka. Denna spricka, eller ”spricka”, kallas klyvningsfåra. Furan fördjupas när aktinringen drar ihop sig, och till slut klyvs membranet i två delar.
I växtceller måste en ny cellvägg bildas mellan dottercellerna. Under interfasen ackumulerar Golgiapparaten enzymer, strukturella proteiner och glukosmolekyler innan den bryts till vesiklar och sprids i hela den delande cellen (figur 12). Under telofasen transporteras dessa Golgivesiklar på mikrotubuli för att bilda en phragmoplast (en vesikulär struktur) vid metafasplattan. Där smälter vesiklarna samman och växer samman från centrum mot cellväggarna; denna struktur kallas cellplatta. När fler vesiklar smälter samman blir cellplattan större tills den smälter samman med cellväggarna i cellens periferi. Enzymer använder den glukos som har ansamlats mellan membranlagren för att bygga en ny cellvägg. Golgimembranen blir delar av plasmamembranet på vardera sidan av den nya cellväggen.
Sammanfattning av mitos och cytokinesi
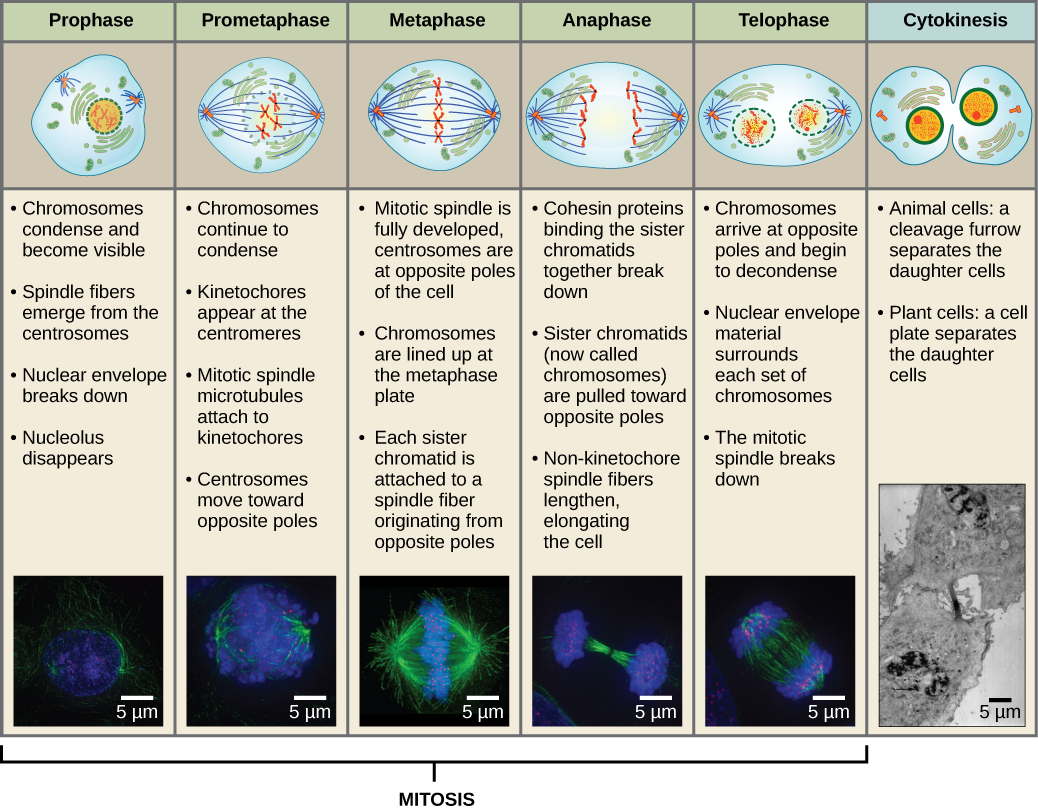
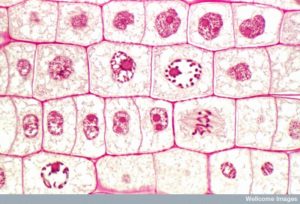
Figur 13 Mitosen är uppdelad i fem stadier – profas, prometafas, metafas, anafas och telofas. Bilderna längst ner har tagits med fluorescensmikroskopi av celler som artificiellt färgats med fluorescerande färgämnen: blå fluorescens visar DNA (kromosomer) och grön fluorescens visar mikrotubuli (spindelapparaten). (kredit ”mitosis drawings”: ändring av Mariana Ruiz Villareal; kredit ”micrographs”: ändring av Roy van Heesbeen; kredit ”cytokinesis micrographs”: Wadsworth Center/New York State Department of Health; scale-bar data från Matt Russell)
G0 Fas
Inte alla celler följer det klassiska cellcykelmönstret där en nybildad dottercell omedelbart går in i interfasen, tätt följt av mitosfasen. Celler i G0-fasen förbereder sig inte aktivt för att dela sig. Cellen befinner sig i ett vilande (inaktivt) stadium och har lämnat cellcykeln. Vissa celler går tillfälligt in i G0-fasen tills en extern signal utlöser att G1-fasen inleds. Andra celler som aldrig eller sällan delar sig, t.ex. mogna hjärtmuskel- och nervceller, förblir i G0 permanent).