Evolution och naturligt urval
Naturen uppmuntrar ingen lättja, förlåter inga misstag
– Ralph Waldo Emerson
Jag har kallat denna princip, enligt vilken varje liten variation, om den är användbar, bevaras, för naturligt urval.
– Charles Darwin, Arternas uppkomst
|
10 okt 2010 |
|
I den här lektionen vill vi ställa följande frågor:
- Hur ledde observationer i naturen till formuleringen av evolutionsteorin?
- Vilka är huvudpunkterna i Darwins evolutionsteori?
- Hur fungerar det naturliga urvalet?
- Vilka bevis har vi för lokal anpassning?
- Hur kan det naturliga urvalet påverka frekvensen av egenskaper under på varandra följande generationer?
Teorins (R)utveckling
Evolutionsteorin är en av de stora intellektuella revolutionerna i mänsklighetens historia och förändrar drastiskt vår uppfattning om världen och vår plats i den. Charles Darwin lade fram en sammanhängande evolutionsteori och samlade en stor mängd bevis till stöd för denna teori. På Darwins tid trodde de flesta vetenskapsmän helt och hållet att varje organism och varje anpassning var skapelsens verk. Linneaus upprättade det system för biologisk klassificering som vi använder i dag, och han gjorde det i en anda av att katalogisera Guds skapelser.
Med andra ord, alla de likheter och olikheter mellan grupper av organismer som är resultatet av den förgreningsprocess som skapar livets stora träd (se figur 1), betraktades av filosofer och vetenskapsmän från det tidiga 1800-talet som en följd av en allsmäktig design.
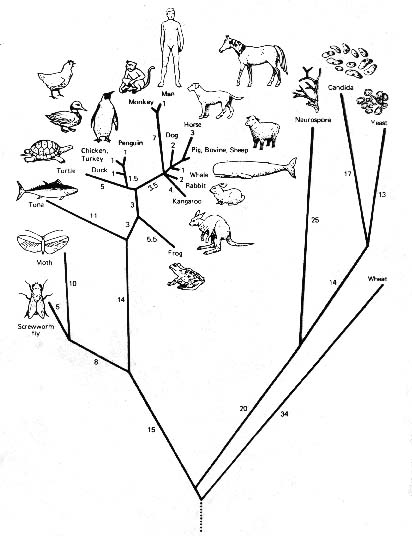
Figur 1: Ett fylogenetiskt ”livets träd” som konstruerats genom datoranalys av cyokrom c-molekyler i de organismer som visas; det finns lika många olika livsträd som det finns analysmetoder för att konstruera dem.
Hursomhelst började ett antal naturhistoriker på 1800-talet att tänka på evolutionär förändring som en förklaring till mönster som observeras i naturen. Följande idéer var en del av det intellektuella klimatet på Darwins tid.
- Ingen visste hur gammal jorden var, men geologer började göra uppskattningar om att jorden var betydligt äldre än vad som förklaras av den bibliska skapelsen. Geologerna lärde sig mer om strata, eller lager som bildas av successiva perioder av avlagring av sediment. Detta tydde på en tidsföljd, där yngre lager överlagrade äldre lager.
- Ett koncept som kallas uniformitarism, som till stor del beror på den inflytelserika geologen Charles Lyell, åtog sig att tyda jordens historia enligt arbetshypotesen att nuvarande förhållanden och processer är nyckeln till det förflutna, genom att undersöka pågående, observerbara processer som erosion och avlagring av sediment.
- Upptäckterna av fossiler ackumulerades under 1700- och 1800-talen. Först trodde naturforskare att de hittade rester av okända men fortfarande levande arter. I takt med att fossilfynden fortsatte blev det dock uppenbart att man inte kände till något som liknade jättedinosaurier från någonstans på planeten. Dessutom påpekade Cuvier redan år 1800 att ju djupare lagren var, desto mindre liknade fossilerna befintliga arter.
- Likheter mellan grupper av organismer ansågs vara bevis på släktskap, vilket i sin tur tydde på evolutionär förändring. Darwins intellektuella föregångare accepterade idén om evolutionära relationer mellan organismer, men de kunde inte ge en tillfredsställande förklaring till hur evolutionen skedde.
- Lamarck är den mest kända av dessa. År 1801 föreslog han organisk evolution som förklaring till den fysiska likheten mellan grupper av organismer och föreslog en mekanism för adaptiv förändring baserad på nedärvning av förvärvade egenskaper. Han skrev om giraffen:
”Vi vet att detta djur, det högsta av däggdjuren, lever i det inre av Afrika, på platser där marken, som nästan alltid är torr och utan örter, tvingar det att söka sig till träden och ständigt anstränga sig för att nå dem. Denna vana, som upprätthållits under lång tid, har hos alla medlemmar av dess ras lett till att frambenen har blivit längre än bakbenen och att halsen har blivit så utsträckt att giraffen, utan att stå på bakbenen, lyfter huvudet till en höjd av sex meter.”
I huvudsak säger detta att giraffernas halsar blev långa som ett resultat av att de hela tiden sträckte sig för att nå höga lövverk. Larmarck hade naturligtvis fel i den antagna mekanismen, men hans exempel visar tydligt att naturforskare tänkte på möjligheten av evolutionär förändring i början av 1800-talet.
- Darwin påverkades av observationer som han gjorde under sin ungdomsresa som naturforskare på undersökningsfartyget Beagle. På Galapagosöarna lade han märke till de små variationer som gjorde att sköldpaddor från olika öar gick att känna igen. Han observerade också en hel rad unika finkar, de berömda ”Darwins finkar”, som uppvisade små skillnader från ö till ö. Dessutom verkade de alla likna, men skilja sig från den vanliga finken på Ecuadors fastland, 600 mil österut. Mönster i organismers utbredning och likhet hade ett viktigt inflytande på Darwins tänkande. Bilden högst upp på den här sidan är Darwins egna skisser av finkar i hans Journal of Researches.
- 1859 publicerade Darwin sin berömda bok On the Origin of Species by Means of Natural Selection (Om arternas uppkomst med hjälp av naturligt urval), ett omfång på över 500 sidor där han samlade in omfattande bevis för sin teori. Publiceringen av boken orsakade ett raseri – varje exemplar av boken såldes samma dag som den släpptes. Medlemmar av det religiösa samfundet, liksom vissa vetenskapliga kollegor, var upprörda över Darwins idéer och protesterade. De flesta vetenskapsmän erkände dock styrkan i Darwins argument. I dag debatterar skolstyrelser fortfarande giltigheten och lämpligheten av Darwins teori i de naturvetenskapliga läroplanerna, och en hel debatt har vuxit fram kring kontroversen (se WWW-webbplatsen Talk.Origins för en pågående dialog). Vi har inte tid att ta upp alla Darwins bevis och argument, men vi kan undersöka de centrala idéerna. Vad säger den här evolutionsteorin?
Darwins teori
Darwins evolutionsteori innebär följande grundläggande idéer. De tre första idéerna diskuterades redan bland tidigare och samtida naturvetare som arbetade med ”artproblemet” när Darwin inledde sin forskning. Darwins ursprungliga bidrag var mekanismen för naturligt urval och rikliga mängder bevis för evolutionär förändring från många källor. Han gav också genomtänkta förklaringar av evolutionens konsekvenser för vår förståelse av livets historia och den moderna biologiska mångfalden.
-
Species (populationer av korsande organismer) förändras över tid och rum. Representanterna för de arter som lever i dag skiljer sig från dem som levde under den senaste tiden, och populationer i olika geografiska områden i dag skiljer sig något i form eller beteende. Dessa skillnader sträcker sig in i fossilregistret, vilket ger gott stöd för detta påstående.
-
Alla organismer har gemensamma förfäder med andra organismer. Med tiden kan populationer dela upp sig i olika arter, som delar en gemensam förfäderspopulation. Långt nog tillbaka i tiden delar alla par av organismer en gemensam förfader. Människor delar till exempel en gemensam förfader med schimpanser för cirka åtta miljoner år sedan, med valar för cirka 60 miljoner år sedan och med kängurur för över 100 miljoner år sedan. Gemensam härstamning förklarar likheterna mellan organismer som klassificeras tillsammans: deras likheter återspeglar arvet av egenskaper från en gemensam förfader.
-
Evolutionär förändring är gradvis och långsam enligt Darwins uppfattning. Detta påstående stöddes av de långa episoderna av gradvis förändring hos organismer i fossilregistret och det faktum att ingen naturforskare hade observerat det plötsliga uppkomsten av en ny art på Darwins tid. Sedan dess har biologer och paleontologer dokumenterat ett brett spektrum av långsamma till snabba evolutionära förändringar inom släkten.
Den primära mekanismen för förändring över tid är det naturliga urvalet, som utvecklas nedan. Denna mekanism orsakar förändringar i egenskaperna (drag) hos organismer inom linjerna från generation till generation.
Processen för naturligt urval
Darwins process för naturligt urval har fyra komponenter.
- Variation. Organismer (inom populationer) uppvisar individuell variation i utseende och beteende. Dessa variationer kan gälla kroppsstorlek, hårfärg, ansiktsmarkeringar, röstegenskaper eller antal avkommor. Å andra sidan uppvisar vissa egenskaper liten eller ingen variation mellan individer – till exempel antalet ögon hos ryggradsdjur.
- Förmåga till arv. Vissa egenskaper förs konsekvent vidare från förälder till avkomma. Sådana egenskaper är ärftliga, medan andra egenskaper påverkas starkt av miljöförhållanden och uppvisar en svag ärftlighet.
- Hög populationstillväxt. De flesta populationer har fler avkommor varje år än vad de lokala resurserna kan försörja vilket leder till en kamp om resurserna. Varje generation upplever en betydande dödlighet.
- Differentierad överlevnad och reproduktion. Individer som har egenskaper som är väl lämpade för kampen om lokala resurser bidrar med fler avkommor till nästa generation.
Från en generation till nästa kommer kampen om resurser (det som Darwin kallade ”kampen för tillvaron”) att gynna individer med vissa variationer framför andra och därmed förändra frekvensen av egenskaper inom populationen. Denna process är det naturliga urvalet. De egenskaper som ger en fördel till de individer som lämnar fler avkommor kallas anpassningar.
För att det naturliga urvalet ska verka på en egenskap måste egenskapen ha en ärftlig variation och ge en fördel i konkurrensen om resurser. Om något av dessa krav inte föreligger, upplever egenskapen inget naturligt urval. (Vi vet nu att sådana egenskaper kan förändras genom andra evolutionära mekanismer som har upptäckts sedan Darwins tid).
Naturligt urval fungerar genom komparativa fördelar, inte genom en absolut standard för design. ”…eftersom det naturliga urvalet verkar genom konkurrens om resurser, anpassar det invånarna i varje land endast i förhållande till graden av perfektion hos deras kompanjoner” (Charles Darwin, On the Origin of Species, 1859).
Under 1900-talet integrerades genetiken med Darwins mekanism, vilket gör det möjligt för oss att utvärdera det naturliga urvalet som den differentiella överlevnaden och reproduktionen av genotyper, som motsvarar särskilda fenotyper. Det naturliga urvalet kan endast verka på befintlig variation inom en population. Sådana variationer uppstår genom mutation, en förändring i någon del av den genetiska koden för en egenskap. Mutationer uppstår av en slump och utan att man förutser den potentiella fördelen eller nackdelen med mutationen. Med andra ord uppstår variationer inte för att de behövs.
Bevis för naturligt urval
Låt oss titta på ett exempel för att göra det naturliga urvalet tydligt.
Industriell melanism är ett fenomen som drabbade över 70 arter av nattfjärilar i England. Det har studerats bäst hos den pepprade nattfjärilen Biston betularia. Före 1800 hade det typiska malet för arten ett ljust mönster (se figur 2). Mörkfärgade eller melaniska malar var sällsynta och var därför samlarobjekt.

Figur 2. Bild av pepparmot
Under den industriella revolutionen gjorde sot och annat industriellt avfall trädstammar mörka och dödade lavar. Den ljusa varianten av malet blev sällsynt och den mörka varianten blev rikligt förekommande. År 1819 sågs den första melaniska varianten. 1886 var den mycket vanligare, vilket visar på en snabb evolutionär förändring. 
Till slut var ljusa morfer vanliga på endast ett fåtal platser, långt från industriområden. Orsaken till denna förändring ansågs vara selektiv predation av fåglar, som gynnade kamouflagefärgningen hos nattfjärilen.
På 1950-talet gjorde biologen Kettlewell försök med återfångst med båda morferna. En kort sammanfattning av hans resultat visas nedan. Genom att observera fågelpredation från persienner kunde han bekräfta att malens uppseendeväckande egenskaper i hög grad påverkade chansen att den skulle bli uppäten.
Framgång vid återfångst
|
ljus nattfjäril |
mörk nattfjäril |
|
|
non-industriella skogar |
14.6 % |
4.7 % |
|
industriell skog |
13 % |
27.5 % |
Lokal anpassning – fler exempel
So långt i dagens föreläsning har vi betonat att det naturliga urvalet är hörnstenen i evolutionsteorin. Det tillhandahåller mekanismen för adaptiv förändring. Varje förändring i miljön (t.ex. en förändring av bakgrundsfärgen på den trädstam som du sitter på) leder sannolikt till lokal anpassning. En utbredd population kommer sannolikt att uppleva olika miljöförhållanden i olika delar av sitt utbredningsområde. Därför kommer den snart att bestå av ett antal delpopulationer som skiljer sig något, eller till och med mycket, från varandra.
Nedan följer exempel som illustrerar populationers anpassning till lokala förhållanden.
- Råttormen, Elaphe obsoleta, har märkbart olika populationer på olika platser i östra Nordamerika (se figur 3). Huruvida dessa ska kallas geografiska ”raser” eller underarter kan diskuteras. Dessa populationer utgör alla en art, eftersom parning kan ske mellan intilliggande populationer, vilket gör att arterna delar en gemensam genpool (se föreläsningen om artbildning).
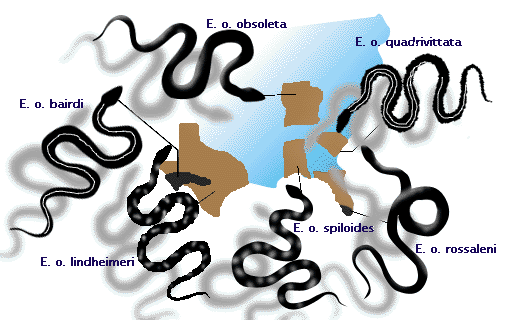
Figur 3: Underarter av råttormen Elaphe obsoleta, som korsar varandra där deras utbredningsområden möts.
- Galapagosfinkarna är det berömda exemplet från Darwins resa. Varje ö på Galapagos som Darwin besökte hade sin egen art av finkar (14 stycken totalt), som inte finns någon annanstans i världen. Vissa hade näbbar som var anpassade för att äta stora frön, andra för små frön, vissa hade papegojliknande näbbar för att äta knoppar och frukter och andra hade smala näbbar för att äta små insekter (se figur 4). En av dem använde en tagg för att söka efter insektslarver i trä, precis som vissa hackspettar gör. (Sex var marklevande och åtta var trädfinkar.) (Denna diversifiering till olika ekologiska roller, eller nischer, anses vara nödvändig för att möjliggöra samexistens av flera arter, ett ämne som vi kommer att undersöka i en senare föreläsning.) För Darwin verkade det som om var och en av dem var något modifierad från en ursprunglig kolonist, troligen finken på Sydamerikas fastland, cirka 600 mil österut. Det är troligt att adaptiv strålning ledde till att så många arter bildades eftersom andra fåglar var få eller saknades, vilket lämnade tomma nischer att fylla, och eftersom Galapagosöarnas många öar gav goda möjligheter till geografisk isolering.
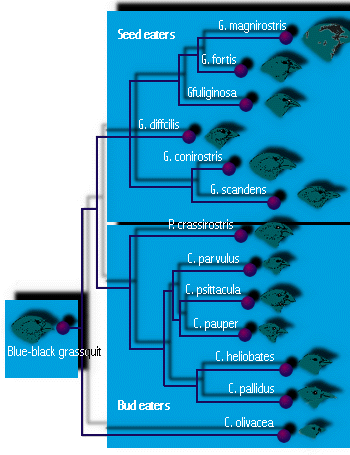
Figur 4
Stabiliserande, riktningsbestämd och diversifierande selektion
Slutligt kommer vi att titta på ett statistiskt sätt att tänka på selektion. Anta att varje population kan beskrivas som en frekvensfördelning för någon egenskap — näbbstorlek, till exempel. Observera återigen att variation i en egenskap är det kritiska råmaterialet för att evolution ska kunna ske.
Hur kommer frekvensfördelningen att se ut i nästa generation?
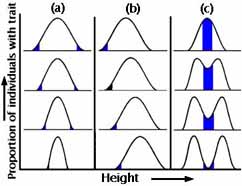
Figurer 5a-c
För det första kan andelen individer med varje värde av egenskapen (näbbstorlek eller kroppsvikt) vara exakt densamma. För det andra kan det finnas en riktningsförändring i bara en riktning. För det tredje (och med sådan sällsynthet att dess existens är diskutabel) kan det finnas en samtidig förändring i båda riktningarna (t.ex. gynnas både större och mindre näbbar, på bekostnad av dem av mellanstorlek). Figurerna 5a-c visar dessa tre huvudkategorier av naturligt urval.
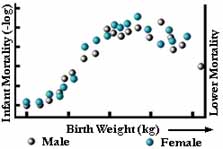
Figur 6
Under stabiliserande urval elimineras extrema sorter från frekvensfördelningens båda ändar. Frekvensfördelningen ser exakt likadan ut som den gjorde i generationen före (se figur 5a). Sannolikt är detta den vanligaste formen av naturligt urval, och vi förväxlar det ofta med inget urval. Ett exempel från verkligheten är födelsevikten hos människobarn (se figur 6).
Under riktat urval klarar sig individer i ena änden av fördelningen av näbbstorlekar särskilt bra, och därför förskjuts frekvensfördelningen av egenskapen i den efterföljande generationen från den plats där den var i föräldragenerationen (se figur 5b). Detta är vad vi vanligtvis tänker på som naturligt urval. Industriell melanism var ett sådant exempel.

Figur 7
Hästens fossila släktlinje ger en anmärkningsvärd demonstration av riktad succession. Den fullständiga härstamningen är ganska komplicerad och är inte bara en enkel linje från den lilla gryningshästen Hyracotherium från tidig eocen till dagens välkända Equus. På det hela taget har hästen dock utvecklats från en liten förfader som är byggd för att röra sig i skogar och buskar till en långbent ättling som är byggd för snabbhet på öppna gräsmarker. Denna utveckling har inneburit väldokumenterade förändringar i tänder, benlängd och tåstruktur (se figur 7).
Under diversifierande (störande) urval gynnas båda extremerna på bekostnad av mellanliggande sorter (se figur 5c). Detta är ovanligt, men av teoretiskt intresse eftersom det antyder en mekanism för artbildning utan geografisk isolering (se föreläsningen om artbildning).
Sammanfattning
Darwins evolutionsteori förändrade i grunden riktningen för framtida vetenskapligt tänkande, även om den byggde på ett växande tankegods som började ifrågasätta tidigare idéer om naturvärlden.
Kärnan i Darwins teori är det naturliga urvalet, en process som sker över successiva generationer och definieras som differentiell reproduktion av genotyper.
Naturligt urval kräver ärftlig variation i en viss egenskap och differentiell överlevnad och reproduktion i samband med innehav av denna egenskap.
Exempel på naturligt urval är väldokumenterade, både genom observation och genom fossilregistret.
Selektion verkar på frekvensen av egenskaper och kan ta formen av stabiliserande, riktat eller diversifierande urval.
Förslag till läsning
o Darwin, C. 1959. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life. London: J. Murray.
o Futuyma, D.J. 1986. Evolutionsbiologi. Sunderland, Mass: Sinauer Associates, Inc.
o Dawkins, R. 1989. The Selfish Gene (Den själviska genen). Oxford: Oxford University Press.