Paranötindustrin — Förr, nu och i framtiden
Scott A. Mori
The New York Botanical Garden
Den här artikeln får återges med tillstånd från Sustainable Harvest and Marketing of Rain Forest Products. Plotkin, M. och L. Famolare (red.). Copyright Island Press, 1992. Publicerad av Island Press, Washington, D.C. & Covelo, Kalifornien. För att beställa ett exemplar av denna bok, ring 1-800-828-1302, eller skriv till: Island Press, Box 7, Covelo, CA 95428
Introduktion | Naturhistoria | Fenologi | Pollineringsbiologi | Spridningsbiologi | Skörd av paranötter |
Plantationer av paranötter | Framtida produktion av paranötter | Referenser
Introduktion
Amazoniens skogar hyser ett stort antal växter med ekonomiskt värde (Balick, 1985; Farnsworth, 1984). Faktum är att intakta Amazonasskogar ofta är mer värdefulla för sina timmer- och andra produkter än de jordbruksplantager eller betesmarker som alltför ofta ersätter dem (Peters et al., 1989; Menezes, 1990). Amazonas skogar har dessutom ett oskattbart värde som reservoar för biologisk mångfald, reglerare av hydrologiska cykler, beskyddare av känsliga jordar och stabilisatorer av atmosfären. På grund av värdet av Amazonas skogar bör de som vill ersätta dem med jordbruk eller betesmarker vara tvungna att visa att deras projekt kommer att ge mer än värdet av de intakta skogarna.
En av de viktigaste ekonomiska växterna i Amazonas är paranöt (Bertholletia excelsa, familjen Lecythidaceae). De ätliga fröna från denna art, tillsammans med latexen från Hevea brasiliensis, nämns ofta som de viktigaste produkterna från utvinningsreserver i Amazonas. Paranötter samlas in främst under den våta säsongen och gummi utvinns främst under den torra säsongen. Kombinationen av dessa två skogsprodukter ger inkomster året runt för dem som lever av extraktivism. Insamlingen av paranötter och gummi har relativt liten inverkan på ekologin i Amazonas skogar. Därför sägs det ofta att bevarandet av den biologiska mångfalden och utnyttjandet av dessa och andra icke-trävaruprodukter är förenliga. De som samlar in andra skogsprodukter än timmer är dock nästan alltid involverade i andra verksamheter, t.ex. svedjebruk  jordbruk, timmerutvinning, gruvdrift och jakt. Som ett resultat av detta skyddar kraftigt utnyttjade utvinningsreservat vanligtvis bara en del av den enorma biologiska mångfald som finns i Amazonas skogar. Följaktligen måste alla bevarandeplaner för Amazonas omfatta stora reserver av alla Amazonas ekosystem som är skyddade från överdriven ekonomisk exploatering.
jordbruk, timmerutvinning, gruvdrift och jakt. Som ett resultat av detta skyddar kraftigt utnyttjade utvinningsreservat vanligtvis bara en del av den enorma biologiska mångfald som finns i Amazonas skogar. Följaktligen måste alla bevarandeplaner för Amazonas omfatta stora reserver av alla Amazonas ekosystem som är skyddade från överdriven ekonomisk exploatering.
I det här kapitlet går jag igenom den naturliga historien, värdet av paranötsskörden, möjligheterna till plantageodling och framtiden för paranötsindustrin. På grund av sin ekonomiska betydelse har paranöten varit föremål för många studier av dess biologi och agronomi. De flesta av dessa studier har utförts under överinseende av ”Centro de Pesquisa Agropecuária do Trópico Umido” (CPATU) vid ”Empresa Brasileira de Pesquisas Agropecuária” (EMBPRAPA) i Belém, Pará, Brasilien. En bibliografi om paranöt med 259 titlar finns tillgänglig (Vaz Pereira och Lima Costa, 1981), och en färsk sammanfattning av paranötets biologi och agronomi finns i Mori och Prance (1990b).
Taxonomi och utbredning
Bertholletia excelsa tillhör en pantropisk trädfamilj (Lecythidaceae) som omfattar cirka 200 arter i Neotropien, fördelade från södra Mexiko till södra Brasilien (Mori och Prance, 1990a; Prance och Mori, 1979). Paranötterna utgör en enda art i det väldefinierade släktet Bertholletia. Även om det finns en avsevärd variation i fruktstorlek och fruktform samt i antalet frön per frukt finns det ingen anledning att erkänna mer än en art av Bertholletia.
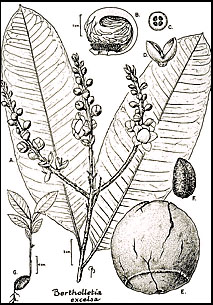 De närmaste släktingarna till B. excelsa finns bland arter av Lecythis som i folkmun brukar kallas jarana-gruppen (Mori och Prance, 1990b). Andra arter med ätliga frön i familjen är Lecythis pisonis och dess släktingar L. minor och L. ollaria. De två sistnämnda arterna orsakar dock ibland hår- och nagelförlust på grund av överdriven selenansamling i fröna (Dickson, 1969; Kerdel-Vegas, 1966). Skillnaderna mellan Bertholletia och Lecythis är dock så stora att det finns små förhoppningar om att införa genetiskt material från det ena släktet till det andra via hybridisering. Följaktligen måste bakterieplasma för att förbättra produktionen av paranötter komma från den variation som finns inom B. excelsa, inte från närbesläktade arter i andra släkten.
De närmaste släktingarna till B. excelsa finns bland arter av Lecythis som i folkmun brukar kallas jarana-gruppen (Mori och Prance, 1990b). Andra arter med ätliga frön i familjen är Lecythis pisonis och dess släktingar L. minor och L. ollaria. De två sistnämnda arterna orsakar dock ibland hår- och nagelförlust på grund av överdriven selenansamling i fröna (Dickson, 1969; Kerdel-Vegas, 1966). Skillnaderna mellan Bertholletia och Lecythis är dock så stora att det finns små förhoppningar om att införa genetiskt material från det ena släktet till det andra via hybridisering. Följaktligen måste bakterieplasma för att förbättra produktionen av paranötter komma från den variation som finns inom B. excelsa, inte från närbesläktade arter i andra släkten.
Bertholletia excelsa är en Amazonasväxt som föredrar icke översvämmad skog (terra firma) i Guianas, Colombia, Venezuela, Peru, Bolivia och Brasilien. De klimatförhållanden under vilka paranötter växer sammanfattas i de Almeida (1963), Diniz och Bastos (1974) samt Mori och Prance (1990b).
Paranötter odlas i tropiska botaniska trädgårdar långt utanför dess naturliga utbredningsområde, och mindre plantager har etablerats i Kuala Lumpur i Malaysia (Müller, 1981) och Ghana i Afrika (D.K. Abbiw, pers. komm.).
Träd av Bertholletia excelsa förekommer i bestånd med 50 till 100 individer som är kända som ”manchales” i Peru (Sánchez, 1973) och ”castanhais” i Brasilien (Dias, 1959). Tätheten av paranötsträd per hektar varierar avsevärt i hela Amazonas. I en studie av paranötsproduktion i östra Brasilien fann Miller (1990) mellan 9 och 26 reproduktiva träd per hektar, medan Becker och Mori (opublicerade uppgifter) fann endast ett träd över 10 centimeter dbh i en 100 hektar stor tomt i centrala Amazonas i Brasilien.
Det finns vissa bevis för att paranötsträd är gapberoende (Mori och Prance, 1990b). Dessutom har vissa författare föreslagit att bestånd av paranötsträd har sitt ursprung från förcolombianska indianer (Miller, 1990; Mori och Prance, 1990b; Müller et al., 1980). En förståelse för utvecklingen av reproduktiva individer från frön behövs fortfarande innan det är möjligt att hantera paranötter i naturliga bestånd.
Blomning av Bertholletia excelsa sker under den torra säsongen och in i den våta säsongen. Paranötter växer faktiskt naturligt endast i regioner med en torrperiod på tre till fem månader (Müller, 1981). I den östra delen av Amazonas Brasilien börjar blomningen i slutet av regnperioden i september och sträcker sig till februari. Den högsta blomningen inträffar i oktober, november och december (Moritz, 1984).
Till slutet av regnperioden, vanligtvis i juli, börjar bladen på paranötsträd falla. Den nya tillväxten skjuter fram direkt under föregående års blomställningar, och de nya blomställningarna produceras vid toppen av den aktuella tillväxtskjutningen. Ett stort antal blommor produceras dagligen under en relativt lång period. Blommorna öppnar sig mellan 4:30 och 5:00 på morgonen. Ståndarna börjar dock att avlägsna sig från knoppen flera timmar innan blommorna öppnas. Kronbladen och androecia faller på eftermiddagen den dag då blommorna öppnar sig (Mori och Prance, 1990b).
Fruktutvecklingen tar längre tid hos B. excelsa än hos någon annan art av Lecythidaceae. Moritz (1984) uppger att det krävs 15 månader för att frukterna ska utvecklas efter att de har satts. Följaktligen faller paranötsfrukterna mest i januari och februari, under regnperioden. Under naturliga förhållanden tar det 12-18 månader för fröna att gro (Müller, 1981).
 Paranötets blommor är zygomorfa, med ett androceum som är förlängt på ena sidan till en huva som böjs över och sitter tätt ihop med toppen av äggstocken. Dessutom sitter kronbladen fast på androceum (fig. 27-1). Följaktligen kan blommorna endast tas in av bin med stora kroppar som är tillräckligt starka för att bryta upp androceumhuvudet för att få den pollineringsbelöning som tros vara nektar som produceras vid toppen av det hoprullade androceumhuvudet. Bin av släktena Bombus, Centris, Epicharis, Eulaema och Xylocopa har fångats när de besökt paranötsträd (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985). Dessa bin är osociala eller halvsociala och lämpar sig därför inte lätt för manipulation av människor, vilket är fallet med de sociala bin (till exempel Apis, Melipona och Trigona) som kan användas för att pollinera vissa grödor genom att transportera bikupor från en plantage till en annan.
Paranötets blommor är zygomorfa, med ett androceum som är förlängt på ena sidan till en huva som böjs över och sitter tätt ihop med toppen av äggstocken. Dessutom sitter kronbladen fast på androceum (fig. 27-1). Följaktligen kan blommorna endast tas in av bin med stora kroppar som är tillräckligt starka för att bryta upp androceumhuvudet för att få den pollineringsbelöning som tros vara nektar som produceras vid toppen av det hoprullade androceumhuvudet. Bin av släktena Bombus, Centris, Epicharis, Eulaema och Xylocopa har fångats när de besökt paranötsträd (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985). Dessa bin är osociala eller halvsociala och lämpar sig därför inte lätt för manipulation av människor, vilket är fallet med de sociala bin (till exempel Apis, Melipona och Trigona) som kan användas för att pollinera vissa grödor genom att transportera bikupor från en plantage till en annan.
 För det mesta behövs korspollinering för frösättning hos neotropiska Lecythidaceae. Därför är bin, och i mindre utsträckning fladdermöss, viktiga för pollineringen och den efterföljande frukt- och fröutvecklingen hos Lecythidaceae. Även om det kan förekomma en låg grad av inavel hos Bertholletia excelsa är de flesta fröställningar hos denna art resultatet av korspollinering (Mori och Prance, 1990b). Utvecklingen av självkompatibla linjer av paranöt skulle underlätta plantageodling av denna art genom att eliminera behovet av korspollinering av de svårhanterliga bi-pollinatörerna.
För det mesta behövs korspollinering för frösättning hos neotropiska Lecythidaceae. Därför är bin, och i mindre utsträckning fladdermöss, viktiga för pollineringen och den efterföljande frukt- och fröutvecklingen hos Lecythidaceae. Även om det kan förekomma en låg grad av inavel hos Bertholletia excelsa är de flesta fröställningar hos denna art resultatet av korspollinering (Mori och Prance, 1990b). Utvecklingen av självkompatibla linjer av paranöt skulle underlätta plantageodling av denna art genom att eliminera behovet av korspollinering av de svårhanterliga bi-pollinatörerna.
Bina utanför paranötets naturliga utbredningsområde kan utföra pollinering. Paranötsträd i Ceylon (Macmillan, 1935), Kuala Lumpur och Ghana sätter till exempel frukt. Det är dock inte känt om pollinatörer utanför paranötets naturliga utbredningsområde eller ”ogräsiga” pollinatörer som finns i sekundära skogar är tillräckligt effektiva för att möjliggöra en ekonomiskt livskraftig fruktproduktion.
 Frukterna och fröna hos B. excelsa är unika inom Lecythidaceae. Vid mognad faller de stora, runda, vedartade frukterna till marken med fröna inuti. Fröna, som har ett benigt testa, avlägsnas från kapslarna och sprids av gnagare, särskilt agoutis (Dasyprocta spp.) (Huber, 1910; Miller; 1990; E. Ortiz, pers. komm.). Agoutis och ekorrar kan vara de enda djur som effektivt kan gnaga sig igenom de extremt vedartade perikarpsen. De äter en del av fröna och gömmer andra för senare användning. Följaktligen äts fröna antingen upp och förstörs eller så lämnas de i en bortglömd gömma där de så småningom kan gro 12 till 18 månader senare.
Frukterna och fröna hos B. excelsa är unika inom Lecythidaceae. Vid mognad faller de stora, runda, vedartade frukterna till marken med fröna inuti. Fröna, som har ett benigt testa, avlägsnas från kapslarna och sprids av gnagare, särskilt agoutis (Dasyprocta spp.) (Huber, 1910; Miller; 1990; E. Ortiz, pers. komm.). Agoutis och ekorrar kan vara de enda djur som effektivt kan gnaga sig igenom de extremt vedartade perikarpsen. De äter en del av fröna och gömmer andra för senare användning. Följaktligen äts fröna antingen upp och förstörs eller så lämnas de i en bortglömd gömma där de så småningom kan gro 12 till 18 månader senare.
 Paranötter skördas nästan helt och hållet från vilda träd under en fem till sex månader lång period under regnperioden. Frukterna, som väger mellan 0,5 och 2,5 kilo och innehåller 10 till 25 frön, samlas in omedelbart efter att de fallit för att minimera insekts- och svampangrepp på fröna och för att kontrollera antalet frön som förs bort av djur (Mori och Prance, 1990b). Enligt Miller (1990) varierar antalet kapslar som produceras per träd från 63 till 216. Mer detaljerade beskrivningar av metoderna för skörd av paranötter finns i Almeida (1963), Mori och Prance (1990b) och Souza (1963).
Paranötter skördas nästan helt och hållet från vilda träd under en fem till sex månader lång period under regnperioden. Frukterna, som väger mellan 0,5 och 2,5 kilo och innehåller 10 till 25 frön, samlas in omedelbart efter att de fallit för att minimera insekts- och svampangrepp på fröna och för att kontrollera antalet frön som förs bort av djur (Mori och Prance, 1990b). Enligt Miller (1990) varierar antalet kapslar som produceras per träd från 63 till 216. Mer detaljerade beskrivningar av metoderna för skörd av paranötter finns i Almeida (1963), Mori och Prance (1990b) och Souza (1963).
Insamling av paranötter har en stor inverkan på lokala ekonomier i Amazonas. Tillgängliga siffror ger dock endast ungefärliga uppgifter om den totala produktionen eftersom det är svårt att få fram exakta uppgifter från Amazonas. Den brasilianska produktionen har varierat från 3 557 ton 1944 till 104 487 ton 1970. Sedan 1980 har den årliga produktionen legat runt 40 000 ton (Mori och Prance, 1990b). Tidigare var välfärden i många städer i Amazonas, såsom Puerto Maldonado i Peru (Sánchez, 1973) och Marabá i Brasilien (Dias, 1959), starkt beroende av produktionen av paranötter. År 1986 uppgick det totala värdet av skalade och oskalade paranötsfrön som exporterades enbart från Manaus till 5 773 228 dollar. (Mori och Prance, 1990b). De flesta av fröna skickas till England, Frankrike, USA och Tyskland.
Beräkningar av Miller (1990) har beräknat det primära värdet (pengar som betalas till insamlarna) av paranötsbestånd till 97 dollar per hektar. I detta värde ingår en godtycklig rabatt på 25 procent för att ta hänsyn till frön som finns kvar i bestånden. Det sekundära värdet – med andra ord de pengar som det exporterande företaget får för oskalade nötter av ett amerikanskt importföretag – uppskattades till 175,56 dollar per hektar. Det tertiära värdet – kostnaden för en påse oskalade nötter i en stormarknad i Florida – beräknades till 1059,44 dollar per hektar. Skalade och bearbetade nötter är mycket mer värdefulla. Ansträngningar, som de som Cultural Survival gör, för att placera en del av skalningen och en del av bearbetningen i samlarnas samhällen ger ytterligare incitament för att behålla utvinningsreserver. Under en tioårsperiod verkar utnyttjandet av en skog för produktion av paranötter vara mer lönsamt än att utvinna timmer eller hugga skogen för bete (Miller, 1990).
Paranötsplantager
Metodiken för odling av paranötter i stora plantager har utvecklats av Müller och hans medarbetare vid CPATU, Belém, Brasilien. En engelsk sammanfattning av deras arbete finns i Mori och Prance (1990b). Som tidigare nämnts samlas den största delen av produktionen av paranötter fortfarande in från vilda träd. Plantager håller dock på att utvecklas i olika delar av Amazonas. I januari 1990 besökte jag Fazenda Aruanã, en paranötsplantage som ligger vid kilometer 215 på vägen Manaus/Itacoatiara i delstaten Amazonas, Brasilien. Detta är en 12 000 hektar stor före detta boskapsranch som delvis omvandlades till paranötsplantage 1980. Vid tiden för mitt besök var 318 660 paranötsträd planterade på 3 341 hektar. Dessutom fanns det ett okänt antal individer som växte i närliggande skogar som tillhör Fazenda Aruanã.
Den ursprungliga avsikten på Fazenda Aruanã var att plantera paranötsträd i rutor på 20 gånger 20 meter och låta boskap beta mellan träden. Till en början åt boskapen inte trädens blad, men senare gjorde de det. Därför minskades avståndet till rutor på 10 gånger 10 meter och antalet nötkreatur minskades till 300 djur. Träden i Aruanã-plantagen är resultatet av ympning av högproduktiva kloner från regionen Abufari Amazonas där paranötterna är kända för sina stora frukter och frön. Ett viktigt övervägande vid anläggandet av denna och andra planteringar är fröets ursprung. Moritz (1984) har visat att fruktproduktionen som ett resultat av befruktning mellan träd av samma klon är låg. Baserat på detta koncept rekommenderar Müller (1981) att knoppar för ympning bör erhållas från minst fem olika träd. En fara med att använda så få kloner för ympning är att urvalet för hög fruktavkastning också kan minska plantagens förmåga att motstå framtida angrepp av sjukdomar och insekter.
Rotstocken odlas på Fazenda Aruanã från frö. Fröna gror genom att de mjukas upp i fuktig sand och sedan tas embryot bort genom att fröskalet öppnas längs kanterna. Embryona, som behandlas med ett svampmedel, gror på cirka 20 dagar och plantorna odlas i plastpåsar eller koppar. När plantorna planteras ut på fältet tillsätts cirka 200 gram fosfor i hålet där de planteras. Rotstocken är redo för ympning efter cirka 1,5 till 2 år när de unga träden har blivit väl etablerade på fältet. Man får bättre ympkvistar om man tar bort bladen från grenarna 8-10 dagar innan ympkvistarna tas bort. Detta främjar bildandet av kraftiga knoppar som klarar sig bättre vid ympning. Trettio dagar efter det att ympen har tagits fram ringmärks rotstocken distalt om ympen. Detta gör det möjligt för ympen att gradvis växa ut ur rotstockens grenar. Efter sex år börjar träden producera frukt. Vid tiden för mitt besök hade dock all frukt som producerats fram till dess använts för att odla mer rotstock eller för att producera plantor för försäljning till andra.
Plantagen verkar inte ha några problem till följd av bristen på pollinatörer. Bixa orellana hade planterats för att tillhandahålla pollen som en födokälla för pollinatörer. Dessutom finns det en omfattande naturskog som omger plantagen och slingor med naturlig vegetation genomkorsar plantagen. I slingorna finns många växter som är kända som födokällor för pollinatörer av paranötsträd. Trots att träden var nästan tio år gamla vid den tidpunkten fanns det inga tecken på eller historik av sjukdom.
Ägarna till Fazenda Aruanã har stora förväntningar på denna plantage. Efter 12 år förväntar de sig en avkastning på 4,8 kilo per träd och 480 dollar per hektar. Efter 18 år räknar de med en avkastning på 8,5 kilo per träd och 850 dollar per hektar.
Framtiden för produktion av paranötter
Intills har paranötter huvudsakligen samlats in från vilda träd. På senare år har produktionen av paranötter minskat på grund av avskogning, utflyttning av paranötssamlare till storstadsområden, översvämning av vissa traditionella paranötsbestånd och kanske på grund av störningar av pollinatörer orsakade av bränder under den torra säsongen när paranötsträden blommar (Kitamura och Müller, 1984; Mori och Prance, 1990b). Om de traditionella metoderna för produktion av paranötter ska bibehållas måste mycket stora utvinningsreservat upprättas i områden med hög täthet av paranötsträd.
De som är intresserade av att bevara den tropiska naturen bör dock vara försiktiga med att sätta likhetstecken mellan inrättandet av utvinningsreservat och bevarandet av den biologiska mångfalden i Amazonas. Eftersom paranötssamlare och gummibrukare gör mer än att bara samla paranötter har de ofta en negativ inverkan på mångfalden av växter och djur. Utvinningsreservat kan i själva verket bli lite mer än sekundär vegetation med ekonomiska växter som paranötter och gummiträd utspridda här och där. Inrättandet av utvinningsreservat förnekar därför inte behovet av välplanerade biologiska reservat.
Den framtida framgången för plantager med paranötter är fortfarande öppen för diskussion. Hittills har det inte funnits några exempel på ekonomiskt framgångsrika planteringar av detta träd från Amazonas. Alla försök att odla tropiska träd i plantager som inte naturligt växer i nästan monotypiska bestånd har misslyckats. I sin naturliga livsmiljö är paranötsträd mer eller mindre utspridda i skogen på ungefär samma sätt som gummiträd växer. Den ekonomiska katastrofen med att försöka föra in gummi i plantager inom dess hemområde har dokumenterats väl (Hecht och Cockburn, 1989), och det finns ingen anledning att tro att paranötsplantager i Amazonas inte kommer att gå samma öde till mötes som de amazoniska gummiplantagerna. En noggrann observation av Aruanã-plantagen under det kommande decenniet kan göra det möjligt för oss att avgöra om produktion av paranötter i plantager är ett livskraftigt alternativ till insamling från vilda träd. Om plantagerna är livskraftiga måste naturvårdare vara beredda att bedöma hur plantagerna kommer att påverka bevarandet av utvinningsreservaten.
Slutningsvis är det viktigt att vi inte sätter alltför stora förhoppningar på utvinning av paranötter som ett ekonomiskt bärkraftigt sätt att försörja en ständigt ökande befolkning i Amazonas. För det första kanske världsmarknaderna inte kan hantera en stor ökning av produktionen av paranötter, och för det andra kan en sådan lågintensiv markanvändning inte försörja en mänsklig befolkning på den nivå som krävs för att öka den levnadsstandard som allt fler människor efterfrågar. Om Ewels (1991) uppskattningar att jakt-samlande och skiftesjordbruk endast kan försörja en person per 5 kvadratkilometer respektive en person per kvadratkilometer är korrekta (det finns ingen anledning att tro att de inte är det), så kommer utvinningsreserverna inte att göra mycket för att absorbera befolkningstillväxten i Amazonas länder. Framtiden för utvinningsreservaten och mänsklighetens förmåga att bevara ett representativt urval av den biologiska mångfalden i Neotropien beror på om man lyckas kontrollera befolkningstillväxten och konsumtionen både i och utanför tropikerna (Erhlich och Erhlich, 1990).
de Almeida, C.P. 1963. Castanha do Para: Sua exportação e importância na economia Amazonica. Edições S.I.A. Estudos Brasileiros 19:1-86.
Balick, M.J. 1985. Useful Plants of Amazonia: A Resource of Global Importance. I G.T. Prance och T.E. Lovejoy (red.), Amazonia. New York: Pergamon Press.
Dias, C.V. 1959. Aspectos geográficos do comércio da castanha no médio Tocantins. Revista Brasil. Geogr. 21(4):77-91.
Dickson, J.D. 1969. Anteckningar om förlust av hår och naglar efter intag av Sapucaia-nötter (Lecythis elliptica). Econ. Bot. 23:133-34.
de Diniz, T.D., A. S. Diniz och T. X. Bastos. 1974. Contribuição ao conhecimento do clima tipico da castanha do Brasil. Bol. Técn. IPEAN 64:59-71.
Ewel, J. 1991. Conservation and Agriculture. Tropinet 2(1):1.
Ehrlich, A.H. och P.R. Ehrlich. 1990. Extinction: Life in Peril. I S. Head och R. Heinzman (red.), Lessons of the Rainforest. San Francisco: Sierra Club Books.
Farnsworth, N.R. 1984. Hur kan brunnen vara torr när den är fylld med vatten? Econ. Bot. 38:4-13.
Hecht, S. och A. Cockburn. 1989. The Fate of the Forest. New York: Verso.
Huber, J. 1910. Mattas e madeiras amazônicas. Bol. Mus. Paraense Hist. Nat. 6:91-225.
Kerdel-Vegas, F. 1966. Den hårborttagande och cytotoxiska effekten av Coco De Mono (Lecythis ollaria) och dess förhållande till kronisk selenios. Econ. Bot. 20:187-95.
Kitamura, P.C. och C.H. Müller. 1984. Castanhais nativas de Marabá-PA: Fatores de depredação e bases para a sua preservação. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Documentos 30:1-32.
Macmillan, H.F. 1935. Tropical Planting and Gardening with Special Reference to Ceylon. London: Macmillan.
Menezes, M.A. 1990. (Reservas extrativistas: Por uma reforma agrária ecológica.) Ciência Hoje 11(64):4-6.
Miller, C. 1990. Natural History, Economic Botany, and Germplasm Conservation of the Brazil Nut Tree (Bertholletia excelsa Humb. and Bonpl). Magisteruppsats framlagd vid University of Florida.
Mori, S. A. och G.T. Prance. 1990a. Lecythidaceae–Part II. The Zygomorphic-Flowered New World Genera (Bertholletia, Corythophora, Couratari, Couroupita, Eschweilera och Lecythis). Fl. Neotrop. Monogr. 21(II): 1-376.
Mori, S. A. och G.T. Prance. 1990b. Taxonomy, Ecology, and Economic Botany of the Brazil nut (Bertholletia excelsa Humb. and Bonpl.: Lecythidaceae). Adv. Econ. Bot. 8:130-50.
Moritz, A. 1984. Biologiska studier av paranöt (Bertholletia excelsa H.B.K.). EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Dokument 29:1-82.
Müller, C. H. 1981. Paranötter; agronomiska studier. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Dokument 2: 1-25.
Müller, C. H., I. A. Rodriques, A. A. Müller och N.R.M. Müller. 1980. Paranöt. Forskningsresultat. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Övrigt 2:1-25.
Nelson, B. W., M. L. Absy, E. M. Barbosa och G. T. Prance. 1985. Observationer om blommande besökare hos Bertholletia excelsa H.B.K. och Couratari tenuicarpa A. C. Sm. (Lecythidaceae). Acta Amazonia 15 (1/2):225-34.
Peters, C.M., A. H. Gentry och R.O. Mendelsohn. 1989. Värdering av en regnskog i Amazonas. Nature 339:655-56.
Prance, G.T., och S.A. Mori. 1979. Lecythidaceae–Part I. De aktinomorfa blommande Lecythidaceae från Nya världen (Asteranthos, Gustavia, Grias, Allantoma och Cariniana). Fl. Neotrop. Monogr. 21(1):1-270.
Sánchez. 1973. Exploatering och saluföring av paranötter i Madre de Dios. Jordbruksministeriet, generaldirektoratet för skogsbruk och jakt, rapport nr 20. Lima, Peru.
Souza A. H. 1963. Castanha do Pará: Botanisk, kemisk och teknisk undersökning. Ediçoes S.I.A., Estudos Técnicos 23:1-69.
Vaz Pereira, I.C., och S.L. Lima Costa. 1981. Bibliografia de Castanha-do-Pará (Bertholletia excelsa H.B.K.). EMPRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. Belém, Pará.
Tillbaka till Lecythidaceae Pages Home
Tillbaka till Mori Home
Granskat i februari 2002. Vid problem eller frågor, vänligen kontakta [email protected].