Člověk spí s cirkadiánní (circa = asi, adia = den) periodicitou a biologové zabývající se cirkadiánními rytmy zkoumali řadu otázek týkajících se tohoto denního cyklu. Co se například stane, když je jedincům znemožněno vnímat signály, které mají normálně o noci a dni? Tato otázka byla zodpovězena umístěním dobrovolníků do prostředí (někdy byly použity jeskyně nebo bunkry) bez vnějších signálů o čase (obrázek 28.3). Během pětidenního období aklimatizace, které zahrnovalo sociální interakce, jídlo v obvyklou dobu a časové signály (rádio, televize), subjekty vstávaly a chodily spát v obvyklou dobu a udržovaly 24hodinový rytmus spánku a bdění. Po odstranění těchto podnětů se však subjekty probouzely každý den později a cyklus spánku a bdění se postupně prodloužil na přibližně 28 hodin místo obvyklých 24 hodin. Když se dobrovolníci vrátili do normálního prostředí, 24hodinový cyklus se rychle obnovil. Lidé (a mnozí další živočichové; viz rámečekB) tedy mají vnitřní „hodiny“, které pokračují v činnosti při absenci jakýchkoli vnějších informací o denní době; za těchto podmínek se říká, že hodiny jsou „volně běžící“.
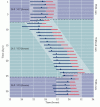
Obrázek 28.3
Rytmus bdění (modré čáry) a spánku (červené čáry) dobrovolníka v izolační komoře s informacemi o denním a nočním cyklu a bez nich.Čísla představují průměr ± směrodatnou odchylku kompletního cyklu bdění/spánku během každého období (více…)
Box B
Molekulární mechanismy biologických hodin.
Předpokládá se, že cirkadiánní hodiny se vyvinuly, aby udržovaly přiměřené doby spánku a bdění navzdory proměnlivému množství denního světla a tmy v různých ročních obdobích a na různých místech planety. K synchronizaci fyziologickýchprocesů s cyklem den-noc (tzv. fotoentréna) musí biologické hodiny detekovat pokles hladiny světla s blížící se nocí. Receptory, které tyto změny světla vnímají, se nepřekvapivě nacházejí ve vnější jaderné vrstvě sítnice, ačkoli odstraněním oka se fotorežim zruší. Detektory však nejsou tyčinky ani čípky. Tyto málo známé buňky leží spíše ve vrstvách gangliových a amakrinních buněk sítnice primátů a myší a promítají se do suprachiasmatického jádra (SCN) hypotalamu, místa cirkadiánní kontroly homeostatických funkcí obecně (obrázek 28.4A). Tyto zvláštní fotoreceptory sítnice obsahují nový fotopigment zvaný melanopsin. Snad nejpřesvědčivějším důkazem roleSCN jako jakýchsi hlavních biologických hodin je skutečnost, že jeho odstranění u pokusných zvířat ruší jejich cirkadiánní rytmus spánku a bdění. SCN řídí i další funkce, které jsou synchronizovány s cyklem spánku a bdění, včetně tělesné teploty (viz obrázek 28.3), vylučování hormonů, produkce moči a změn krevního tlaku. Buněčné mechanismy cirkadiánní regulace jsou shrnuty v rámečkuB.

Obrázek 28.4
Anatomické základy cirkadiánních rytmů. (A) Hypotalamus,ukazující umístění suprachiasmatického jádra (SCN), které je u živočichů primárními „biologickými hodinami“. Název „suprachiasmatické“ je odvozen od (více…)
Aktivace suprachiazmatického jádra vyvolává reakce neuronů, jejichž axonysestupují do pregangliových sympatických neuronů v postranním rohu míchy (obrázek 28.4B). Tyto buňky zase modulují neurony v horních krčních gangliích, jejichž postgangliové axony vystupují do epifýzy (epifýza znamená tvar šišky) ve střední čáře poblíž dorzálního talamu. Epifýza syntetizuje z tryptofanu neurohormonemelatonin (N-acetyl-5-methoxytryptamin) podporující spánek a vylučuje ho do krevního oběhu, aby pomohla modulovat obvody mozkového kmene, které nakonec řídí cyklus spánek-bdění (viz str. 615 a násl.). Je předvídatelné, že syntéza melatoninu se zvyšuje s ubývajícím světlem a dosahuje svého maxima mezi 2:00 a 4:00. U starších lidí šišinka vápenatí a melatoninu se produkuje méně, což možná vysvětluje, proč starší lidé spí méně hodin a častěji trpí nespavostí.
.