Eukaryota mají dva hlavní typy buněčného dělení: mitózu a meiózu. Mitóza slouží k tvorbě nových tělních buněk pro růst a hojení, zatímco meióza k tvorbě pohlavních buněk (vajíček a spermií). O meióze bude pojednáno v pozdější kapitole.
Buněčný cyklus je uspořádaná řada událostí zahrnující buněčný růst a buněčné dělení, při nichž vznikají dvě nové dceřiné buňky prostřednictvím mitózy. Délka buněčného cyklu je velmi variabilní i v rámci buněk jednotlivého organismu. U člověka se frekvence buněčné obměny pohybuje od několika hodin v raném embryonálním vývoji po průměrně dva až pět dní u epiteliálních buněk nebo po celý lidský život strávený bez dělení u specializovaných buněk, jako jsou kortikální neurony nebo buňky srdečního svalu. Doba, kterou buňka stráví v jednotlivých fázích buněčného cyklu, se rovněž liší. Pokud jsou rychle se dělící savčí buňky pěstovány v kultuře (mimo tělo za optimálních podmínek růstu), je délka cyklu přibližně 24 hodin. Načasování událostí v buněčném cyklu je řízeno mechanismy, které jsou jak vnitřní, tak vnější pro buňku.
Buňky na cestě k buněčnému dělení procházejí řadou přesně načasovaných a pečlivě regulovaných fází růstu, replikace DNA a dělení, které vytvářejí dvě geneticky identické buňky. Buněčný cyklus má dvě hlavní fáze: interfázi a mitotickou fázi (obr. 1). Během interfáze buňka roste a dochází k replikaci DNA. Během mitotické fáze se replikovaná DNA a cytoplazmatický obsah oddělí a buňka se rozdělí.
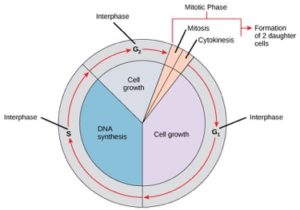
Během interfáze probíhají v buňce běžné procesy a zároveň se připravuje na buněčné dělení. Aby buňka přešla z interfáze do mitotické fáze, musí být splněno mnoho vnitřních i vnějších podmínek. Tři fáze interfáze se nazývají G1, S a G2.
Fáze G1 (první mezera)
První fáze interfáze se nazývá fáze G1 (první mezera), protože z mikroskopického hlediska je vidět jen málo změn. Během fáze G1 je však buňka na biochemické úrovni poměrně aktivní. Buňka hromadí stavební kameny chromozomální DNA a přidružené proteiny a také akumuluje dostatečné zásoby energie k dokončení úkolu replikace každého chromozomu v jádře.
Fáze S (syntéza DNA)
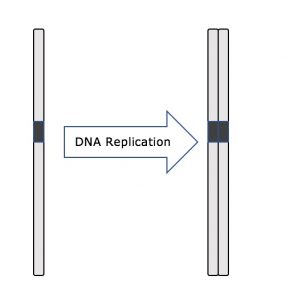
Po celou dobu interfáze zůstává jaderná DNA v polozhu kondenzovaného chromatinu. Ve fázi S může replikace DNA probíhat prostřednictvím mechanismů, které vedou k vytvoření identických párů molekul DNA – sesterských chromatid – pevně spojených s centromerickou oblastí (obr. 2).
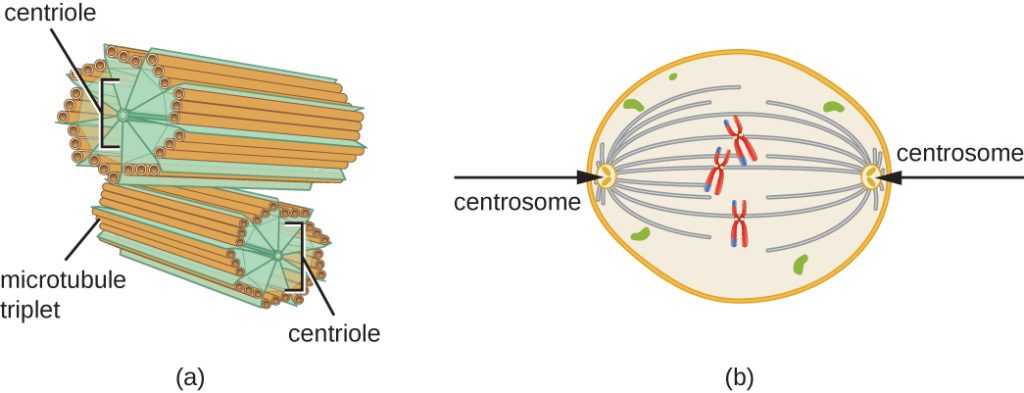
Centrozom se během S fáze rovněž duplikuje. Ze dvou centrozomů vznikne mitotické vřeténko, aparát, který organizuje pohyb chromozomů během mitózy. V centru každé živočišné buňky jsou centrozomy živočišných buněk spojeny s dvojicí tyčinkovitých objektů, centriol, které k sobě svírají pravý úhel. Centrioly pomáhají organizovat buněčné dělení. Centrioly nejsou přítomny v centrozomech jiných eukaryotických druhů, jako jsou rostliny a většina hub.
Fáze G2 (druhá mezera)
Ve fázi G2 buňka doplňuje své energetické zásoby a syntetizuje proteiny nezbytné pro manipulaci s chromozomy. Některé buněčné organely jsou zdvojeny a cytoskelet je demontován, aby poskytl zdroje pro mitotickou fázi. Během G2 může docházet k dalšímu růstu buněk. Než je buňka schopna vstoupit do první fáze mitózy, musí být dokončeny poslední přípravy na mitotickou fázi.
Mitotická fáze
Aby vznikly dvě dceřiné buňky, musí se rozdělit obsah jádra a cytoplazmy. Mitotická fáze je několikastupňový proces, během něhož se zdvojené chromozomy vyrovnají, oddělí a přesunou na opačné póly buňky a poté se buňka rozdělí na dvě nové identické dceřiné buňky. První část mitotické fáze, mitóza, se skládá z pěti fází, které uskutečňují jaderné dělení (obrázek 5). Druhá část mitotické fáze, nazývaná cytokineze, představuje fyzické oddělení cytoplazmatických složek na dvě dceřiné buňky. Ačkoli jsou fáze mitózy u většiny eukaryot podobné, proces cytokineze je zcela odlišný u eukaryot, která mají buněčné stěny, například u rostlinných buněk.
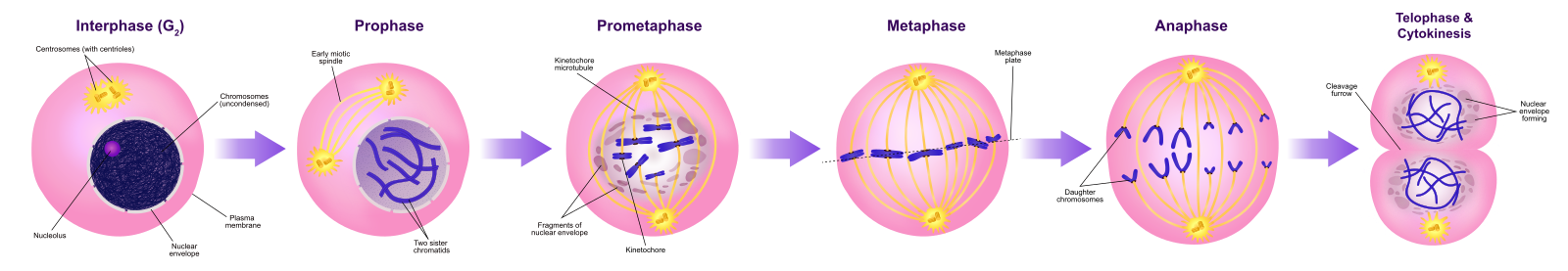
Fáze
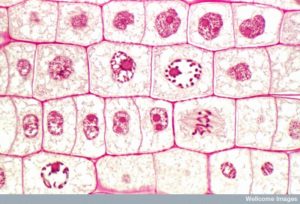
Během profáze, „první fáze“, se jaderný obal začne rozpadat na malé vezikuly a membránové organely (například Golgiho aparát a endoplazmatické retikulum), se fragmentují a rozptylují směrem k okrajům buňky. Jádro zaniká. Centrozomy se začnou přesouvat k opačným pólům buňky. Mikrotubuly, které budou tvořit mitotické vřeténko, se prodlužují mezi centrozomy a tlačí je dál od sebe, jak se vlákna mikrotubulů prodlužují. Sesterské chromatidy se začnou těsněji svinovat pomocí proteinů kondenzinu a stanou se viditelnými pod světelným mikroskopem.
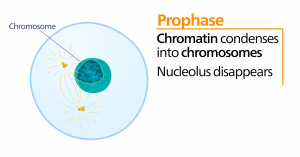
Prometafáze
V průběhu prometafáze, „fáze první změny“, pokračují mnohé procesy, které byly zahájeny v profázi. Fragmentují se zbytky jaderného obalu. Mitotické vřeténko pokračuje ve vývoji, protože se sestavují další mikrotubuly a táhnou se po celé délce bývalé jaderné oblasti. Chromozomy se stávají kondenzovanějšími a diskrétnějšími. U každé sesterské chromatidy vzniká v centromerické oblasti proteinová struktura zvaná kinetochor.
Bílkoviny kinetochoru přitahují a vážou mikrotubuly mitotického vřeténka. Jak se mikrotubuly vřeténka vysouvají z centrozomů, některé z těchto mikrotubulů se dostávají do kontaktu s kinetochory a pevně se na ně vážou. Jakmile se mitotické vlákno připojí k chromozomu, chromozom se orientuje tak, aby kinetochory sesterských chromatid směřovaly k opačným pólům. Nakonec se všechny sesterské chromatidy připojí prostřednictvím svých kinetochorů k mikrotubulům z opačných pólů. Mikrotubuly vřeténka, které nezapojují chromozomy, se nazývají polární mikrotubuly. Tyto mikrotubuly se vzájemně překrývají uprostřed mezi oběma póly a přispívají k prodlužování buňky. Astrální mikrotubuly se nacházejí v blízkosti pólů, napomáhají orientaci vřeténka a jsou nezbytné pro regulaci mitózy.

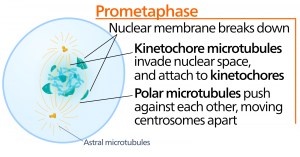
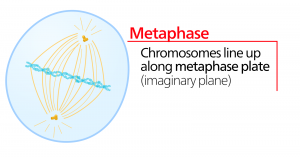
Metafáze
Během metafáze, „fáze změny“, jsou všechny chromozomy zarovnány do roviny zvané metafázová destička nebo rovina rovníku, uprostřed mezi oběma póly buňky. Sesterské chromatidy jsou k sobě stále pevně připojeny pomocí kohesinových proteinů. V této době jsou chromozomy maximálně kondenzované.
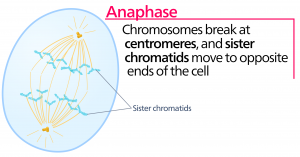
Anafáze
Během anafáze, „vzestupné fáze“, dochází k degradaci kohesinových proteinů a sesterské chromatidy se oddělují v centroméře. Každá chromatida, nyní nazývaná chromozom, je rychle tažena k centrozomu, ke kterému je připojena její mikrotubule. Buňka se viditelně prodlouží (má oválný tvar), protože polární mikrotubuly kloužou proti sobě na metafázní destičce, kde se překrývají.
Telofáze
Během telofáze, „distanční fáze“, dosáhnou chromozomy opačných pólů a začnou se dekondenzovat (rozplétat), čímž se uvolní do chromatinového uspořádání. Mitotická vřeténka jsou depolymerizována na tubulinové monomery, které budou použity k sestavení cytoskeletálních komponent pro každou dceřinou buňku. Kolem chromozomů se vytvoří jaderné obaly a v jaderné oblasti se objeví nukleozomy.
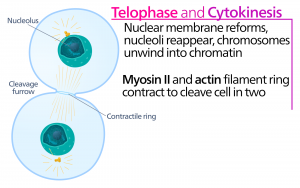
Cytokineze
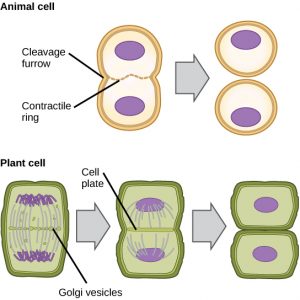
Cytokineze neboli „pohyb buňky“ je druhou hlavní fází mitotické fáze, během níž je dokončeno buněčné dělení prostřednictvím fyzického oddělení cytoplazmatických složek na dvě dceřiné buňky. Dělení není dokončeno, dokud nejsou buněčné složky rozděleny a zcela odděleny na dvě dceřiné buňky. Ačkoli jsou fáze mitózy u většiny eukaryot podobné, proces cytokineze je zcela odlišný u eukaryot, která mají buněčné stěny, například u rostlinných buněk.
U buněk, jako jsou živočišné buňky, které nemají buněčné stěny, následuje cytokineze po nástupu anafáze. Těsně uvnitř plazmatické membrány na bývalé metafázní destičce se vytvoří kontraktilní prstenec složený z aktinových vláken (obr. 12). Aktinová vlákna vtahují rovník buňky dovnitř a vytvářejí trhlinu. Tato trhlina se nazývá štěpná rýha. Brázda se prohlubuje, jak se aktinový prstenec smršťuje, a nakonec se membrána rozštěpí na dvě části.
V rostlinných buňkách se mezi dceřinými buňkami musí vytvořit nová buněčná stěna. Během interfáze se v Golgiho aparátu hromadí enzymy, strukturální proteiny a molekuly glukózy, než se rozpadnou na vezikuly a rozptýlí se po celé dělící se buňce (obr. 12). Během telofáze jsou tyto Golgiho vezikuly transportovány na mikrotubulech, aby vytvořily fragmoplast (vezikulární strukturu) na metafázové destičce. Tam se vezikuly spojí a slijí od středu směrem k buněčným stěnám; tato struktura se nazývá buněčná destička. Jak se spojují další vezikuly, buněčná destička se zvětšuje, až splyne s buněčnými stěnami na periferii buňky. Enzymy využívají glukózu, která se nahromadila mezi vrstvami membrán, ke stavbě nové buněčné stěny. Golgiho membrány se stanou součástí plazmatické membrány na obou stranách nové buněčné stěny.
Souhrn mitózy a cytokineze
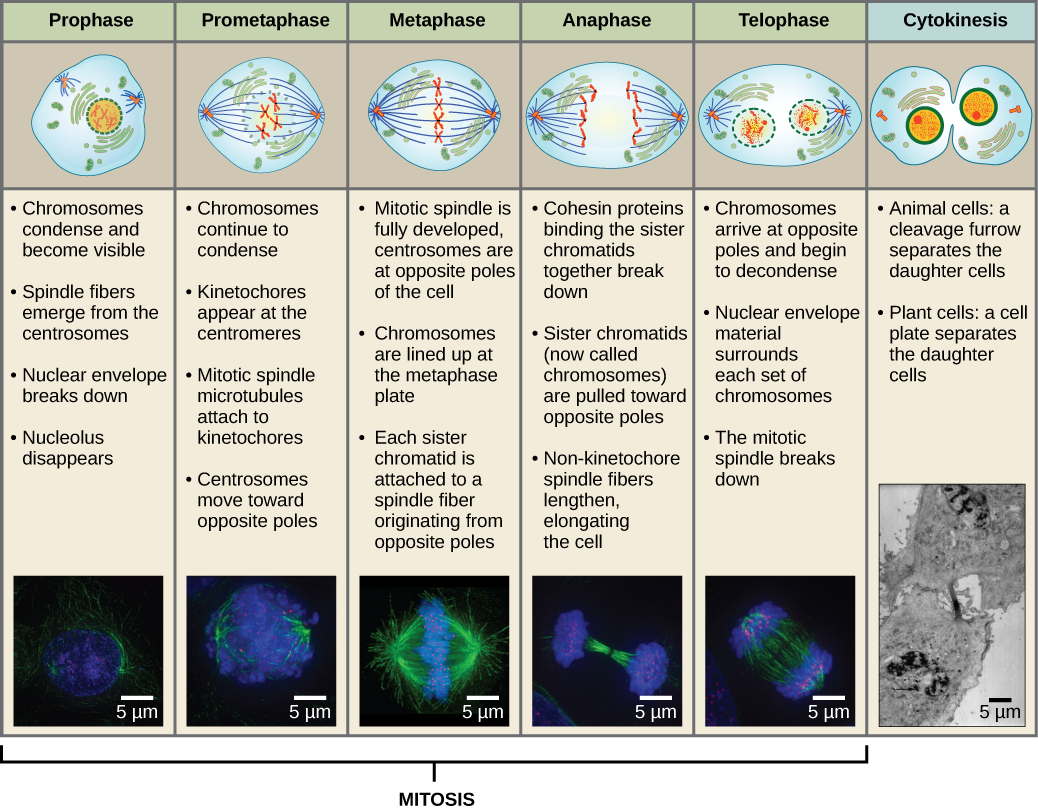
Obrázek 13 Mitóza se dělí na pět fází – profázi, prometafázi, metafázi, anafázi a telofázi. Obrázky v dolní části byly pořízeny fluorescenční mikroskopií buněk uměle obarvených fluorescenčními barvivy: modrá fluorescence označuje DNA (chromozomy) a zelená fluorescence mikrotubuly (vřeténkový aparát). (kredit „mitosis drawings“: úprava práce Mariany Ruiz Villareal; kredit „micrographs“: úprava práce Roye van Heesbeena; kredit „cytokinesis micrograph“: Wadsworth Center/New York State Department of Health; data s měřítkem od Matta Russella)
Fáze G0
Ne všechny buňky dodržují klasické schéma buněčného cyklu, kdy nově vzniklá dceřiná buňka okamžitě vstupuje do interfáze, po níž těsně následuje mitotická fáze. Buňky ve fázi G0 se aktivně nepřipravují na dělení. Buňka se nachází v klidové (neaktivní) fázi a opustila buněčný cyklus. Některé buňky vstupují do fáze G0 dočasně, dokud vnější signál nevyvolá nástup fáze G1. Jiné buňky, které se nikdy nebo jen zřídka dělí, například zralé buňky srdečního svalu a nervové buňky, zůstávají v G0 trvale).
.