Introduzione
Un precedente articolo del nostro laboratorio (Shalom, 2009) sosteneva che l’elaborazione neurocognitiva può essere divisa in tre livelli: (1) un livello di base che coinvolge l’elaborazione cognitiva primaria, emotiva e sensorimotoria. Per esempio, un forte suono inaspettato che viene percepito nei sistemi uditivi-sensoriali primari potrebbe innescare risposte emotive fisiologiche, come la paura (battiti cardiaci veloci, ecc.). (2) un livello integrativo, che combina l’output di tutti i processi primari del livello base e forma un significato, un’esperienza o un comportamento globale e coerente. Per esempio, le rappresentazioni mentali dei vari elementi primari che costituiscono la risposta emotiva di paura si traducono in una sensazione cosciente di avere paura. (3) Un livello logico, che forma regole logiche astratte (regole if-then) dal livello di base (per esempio, “Se ho battiti cardiaci veloci e sudore freddo, allora potrei avere paura”), ed è anche coinvolto nella selezione e nell’inibizione (per esempio, il controllo degli impulsi emotivi). Questa architettura a tre livelli è stata applicata a quattro domini psicologici generali: emozione, memoria, sensazione-percezione e motore. Shalom (2009) si è concentrato sul secondo livello integrativo e sulla sua relazione con i quattro domini psicologici. Ha anche sostenuto che questi quattro tipi di integrazione sono serviti da quattro diverse sottoaree della corteccia prefrontale mediale: Area Brodmann (BA) 11 (percezione), BA 10 (memoria), BA 9 (emozione), e BA 8 (motore). Infine, ha presentato le prove che un deficit selettivo in queste aree BA e questi tipi di processi integrativi sono alla base di alcuni dei deficit comuni in ASD (disturbi dello spettro autistico).
L’attuale revisione tenta di eseguire un’analisi simile per la corteccia prefrontale laterale (LPFC), passando dalle aree prefrontali mediali rilevanti alle loro controparti laterali: BA 11/BA 47 laterale (percezione), BA 10/BA 46 laterale (memoria), BA 9 laterale (emozione) e BA 8 laterale (motore) (Figura 1), (cfr. la distinzione tra cluster 1, 2, 3 e 4 del cingolo anteriore in Beckmann et al. (2009). Porta l’evidenza che un’atipicità selettiva in queste aree BA e un deficit selettivo nei processi di selezione/inibizione in questi quattro domini cognitivi è coinvolto in alcuni dei deficit comuni in ADHD (disturbo da deficit di attenzione e iperattività), come le difficoltà di inibire le risposte motorie (es, l’incapacità di inibire i movimenti inappropriati), l’attenzione percettiva (per esempio, l’incapacità di ignorare la distrazione) e le reazioni emotive (per esempio, l’incapacità di controllare gli impulsi) (Barkley, 1997; Nigg, 1999).
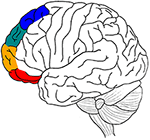
Figura 1. Un riassunto grafico delle principali ipotesi anatomiche dell’articolo. Elaborazione motoria in blu, emozione in verde, memoria in arancione, e sensoriale-percettiva in rosso.
Percezione
Selezione e inibizione percettiva e BA 11/47 laterale
Ci sono prove che la selezione e l’inibizione percettiva sono supportate da reti neurali che coinvolgono la parte laterale della BA 11/47 nella LPFC. In un modello proposto da Zsuga et al. (2016), si suggerisce che la corteccia orbitofrontale (OFC) svolga un ruolo importante nella selezione degli stimoli visivi in base ai requisiti del compito.In particolare, essi suggeriscono che la parte mediale dell’OFC svolge un ruolo centrale nell’integrare e categorizzare le informazioni sugli stimoli dall’ambiente e il loro contesto, mentre la parte laterale dell’OFC è coinvolta nell’assegnare e aggiornare i parametri di selezione in base ai valori specifici del compito degli stimoli disponibili. A sostegno di questa affermazione, in uno studio, Howard e Kahnt (2017) hanno dimostrato che la parte laterale del BA 11 è coinvolta nella codifica dei valori diretti agli obiettivi degli stimoli olfattivi. Hanno presentato dei partecipanti affamati con due odori alimentari preferiti e hanno chiesto loro di scegliere se annusare l’uno o l’altro. Più tardi, i partecipanti sono stati dati uno dei due alimenti da mangiare e sono stati autorizzati a scegliere tra i due odori ancora una volta. Il cambiamento nella preferenza verso l’odore non saziato è stato riflesso da un cambiamento di attività nella parte laterale del BA 11/47. Inoltre, è stato dimostrato che il BA 11/47 laterale è coinvolto nel rifiuto di stimoli irrilevanti (Kaufman et al., 2016).
Il BA 11/47 laterale ha anche dimostrato di essere coinvolto nel mantenimento degli stimoli visivi nella memoria di lavoro e nella guida della selezione visiva. In uno studio, Soto et al. (2007), hanno confrontato due condizioni di una selezione visiva innescata in cui il primo è stato mostrato una o due volte. L’attivazione laterale del BA 11/47 era maggiore quando il primo veniva mostrato due volte, tuttavia, una riduzione dell’attivazione è stata trovata quando stimoli simili venivano ripetuti passivamente due volte (senza una fase di selezione visiva, e quindi senza rilevanza del compito).
Infine, secondo Price (2007), il BA 11 laterale fa parte della rete orbitale sensoriale.
ADHD e selezione e inibizione visiva
Ci sono prove che bambini e adulti con ADHD hanno problemi con l’inibizione di stimoli visivi e uditivi irrilevanti. Per esempio, gli adulti con ADHD hanno mostrato più effetti di interferenza (cioè, problemi nell’inibire gli stimoli irrilevanti) rispetto ai controlli quando è stato chiesto di completare un compito di richiamo ignorando un rumore di fondo (Pelletier et al., 2016). Inoltre, è stata trovata una correlazione positiva tra le prestazioni del compito in condizioni di suono irrilevante e l’entità dei sintomi attenzionali riportati dai pazienti su una scala di sintomi clinici. In un altro studio, agli adulti ADHD e di controllo è stato chiesto di condurre una conversazione telefonica durante la guida. Il gruppo ADHD ha mostrato capacità di guida significativamente più povere durante la condizione di conversazione telefonica che nella condizione di silenzio, se confrontato con i controlli (Reimer et al., 2010). Altri studi hanno dimostrato scarsa inibizione visiva in individui ADHD. Ad esempio, Forster et al. (2014) e Forster e Lavie (2016) hanno scoperto che gli adulti ADHD hanno eseguito un compito di ricerca delle lettere più lentamente quando sullo schermo erano presenti personaggi irrilevanti ma salienti dei cartoni animati, in misura maggiore rispetto ai controlli. In modo correlato, il compito di interferenza Stroop richiede ai partecipanti di frequentare una dimensione specifica di uno stimolo visivo ignorandone un’altra. Gli studi hanno scoperto che i bambini ADHD hanno prestazioni più scarse nella condizione di interferenza Stroop rispetto ai controlli (Sørensen et al., 2014), e che le loro prestazioni sono correlate con i loro sintomi di disattenzione e iperattività (Ikeda et al., 2013).
ADHD, selezione visiva e inibizione e BA 11/47 laterale
Inoltre, diversi studi hanno trovato prove specifiche di attivazione atipica nel BA 11/47 laterale in individui con ADHD che eseguono compiti di selezione/inibizione visiva. Tsujimoto et al. (2013) ha chiesto ai bambini con e senza ADHD di eseguire un compito di memoria di lavoro con e senza distrazione. I partecipanti ADHD hanno mostrato prestazioni comportamentali significativamente più scarse, in particolare sotto distrazione, e hanno mostrato livelli significativamente più elevati di attivazione nel BA 11/47 laterale rispetto ai controlli. In uno studio diverso, Yasumura et al. (2014) hanno testato i bambini ADHD e di controllo su entrambe le condizioni di interferenza Stroop e interferenza Stroop inversa, e ha trovato che in entrambe le condizioni i bambini ADHD hanno eseguito più male e che le prestazioni correlate con l’attività BA 47 laterale. In un altro studio in cui un distrattore saliente ma irrilevante è stato presentato durante un compito di stima del tempo, è stato trovato che mentre il distrattore ha assistito i partecipanti ADHD allo stesso grado dei controlli, l’attivazione BA 11 laterale che ha accompagnato la comparsa del distrattore era significativamente più grande (Pretus et al, 2016).
Memoria
Selezione e inibizione della memoria e BA 10/46 laterale
Mentre non è un consenso generale che il BA10/46 sia impegnato nelle funzioni di memoria episodica, ci sono molte prove a sostegno di un ruolo del BA 10/46 nella selezione e inibizione delle informazioni di memoria. Per esempio, Kim (2011), ha analizzato 74 studi che hanno confrontato il ricordo e la dimenticanza (cioè, elementi ricordati con successo contro distrattori giudicati erroneamente apparsi), e ha trovato che la maggior parte degli studi si è sovrapposta alla parte frontale della BA 46, vicino alla sua giunzione con la BA 10 laterale. In uno studio successivo, Kim (2013) ha eseguito una meta-analisi di 70 studi concentrandosi su elementi nuovi e vecchi (cioè, hit vs. rifiuti corretti). I risultati mostrano che i confronti nuovi>vecchi producono attivazioni che si diffondono nella parte centrale del prefrontale laterale sinistro. Tuttavia, quando ci si concentra su studi di memoria di origine in cui i partecipanti sono tenuti a recuperare non solo l’elemento di destinazione, ma anche un ulteriore dettaglio relativo alla voce dalla fase di apprendimento (cioè, quando le richieste di selezione sono più elevate), l’attivazione specifica è stata vista nella parte frontale del BA46, compresa la parte rostrale del BA10.
Un’altra linea di evidenza proviene da una meta-analisi di Gilbert et al. (2006), in cui sono stati analizzati 104 studi incentrati sul BA 10, e si è scoperto che la maggior parte degli studi che coinvolgono la parte laterale del BA 10 ha testato la memoria di lavoro e il recupero della memoria episodica, mentre gli studi che coinvolgono la parte mediale del BA 10 hanno testato la mentalizzazione (cioè, partecipare alle proprie emozioni e stati mentali o quelli di altri agenti).
Inoltre, Cohen et al. (2014), hanno impiegato il compito di ricordare diretto al valore, in cui i partecipanti sono presentati con elenchi di parole in cui alcune parole sono assegnate più valore di altre, incoraggiando così i partecipanti a tentare e selezionare elementi di valore più elevato rispetto a quelli di basso valore durante la codifica. I risultati mostrano un’attivazione specifica del BA46/10 laterale destro, e un’attivazione molto pronunciata del BA46/10 laterale sinistro, quando appariva un segnale che prediceva una parola di alto valore.
ADHD e selezione e inibizione della memoria
Ci sono diversi studi che hanno trovato prove di difficoltà nella selezione e inibizione delle informazioni di memoria nell’ADHD. Per esempio, in uno studio di Pollak et al. (2007), adulti con e senza ADHD hanno eseguito un difficile compito di apprendimento di liste in cui dovevano memorizzare e ricordare cinque liste di parole in otto diverse occasioni. I risultati mostrano che i partecipanti con ADHD hanno fatto più doppi richiami ed errori di intrusione (ricordando parole dalla lista sbagliata), che sono esempi di una difficoltà nell’inibire il recupero di elementi irrilevanti per il compito. In uno studio simile, Soliman e Elfar (2017), hanno chiesto ad adulti con e senza ADHD di imparare otto liste e sono stati poi presentati con un compito di riconoscimento. Il gruppo ADHD ha riconosciuto meno parole correttamente, ha prodotto più risposte false positive (cioè, selezionando erroneamente elementi che non sono stati appresi) ed erano più sicuri dei loro errori. In un altro studio, Castel et al. (2011), hanno chiesto a bambini con e senza ADHD di eseguire il paradigma del ricordo diretto al valore di cui sopra. I risultati mostrano che i bambini con ADHD hanno ricordato tanti elementi quanti i controlli, tuttavia sono stati meno capaci di selezionare quali elementi ricordare.
ADHD, selezione e inibizione della memoria e BA 10/46 laterale
La ricerca della letteratura ha prodotto solo uno studio di neuroimaging che valuta la selezione o l’inibizione delle funzioni di memoria nell’ADHD. In uno studio di Depue et al. (2010), agli adulti con e senza ADHD è stata insegnata una relazione tra coppie faccia-immagine fino alla padronanza. In seguito è stata mostrata loro l’immagine di un volto ed è stato chiesto loro di pensare all’immagine correlata o di evitare che l’immagine correlata entrasse nella coscienza. I risultati mostrano che i partecipanti ADHD hanno attivato il BA 10 laterale meno dei controlli quando hanno confrontato la condizione “non pensare” con la condizione “pensare”.
Emozione
Selezione e inibizione delle emozioni e BA 9 laterale
Ci sono molte prove a sostegno della relazione tra la regolazione delle emozioni (inibizione di certe emozioni e selezione di altre) e BA 9 laterale. Per esempio, in uno studio di Hallam et al. (2014), i partecipanti hanno visto film che inducono emozioni e hanno dovuto sopprimere o rivalutare la loro reazione emotiva, o semplicemente guardare il film. I risultati hanno mostrato che sia la rivalutazione che la soppressione rispetto alla condizione passiva hanno mostrato l’attivazione del BA 9 laterale. Allo stesso modo, due studi hanno confrontato la rivalutazione e la visione passiva di immagini in persone con PTSD e controlli. Entrambi gli studi hanno trovato che il BA 9 laterale si attivava di più durante la condizione di rivalutazione, ma in misura minore nelle persone con PTSD (Xiong et al., 2013; Rabinak et al., 2014).
Inoltre, due diverse meta-analisi sono state utilizzate per esaminare gli studi fMRI di valutazione della regolazione delle emozioni. Buhle et al. (2014) hanno trovato che la rivalutazione che coinvolge la regolazione verso il basso dell’affetto negativo ha attivato in modo coerente il BA 9 laterale; Frank et al. (2014) hanno trovato che tale rivalutazione era accompagnata da una maggiore attivazione nel BA 9 laterale e una minore attivazione nell’amigdala.
ADHD e selezione e inibizione delle emozioni
Diversi studi hanno trovato deficit di regolazione delle emozioni nei bambini con ADHD. Per esempio, diversi studi mostrano che i bambini con ADHD hanno più difficoltà a sopprimere le loro emozioni rispetto ai loro coetanei con sviluppo tipico (TD). Per esempio, in uno studio i bambini sono stati sottoposti a un compito di competizione tra pari, in cui la metà è stata richiesta di nascondere le loro emozioni e l’altra metà no. Secondo le valutazioni fatte da giudici ingenui, i risultati mostrano che i bambini ADHD erano meno capaci di mascherare le loro emozioni rispetto ai loro coetanei (Walcott e Landau, 2004). Inoltre, uno studio che ha valutato le prestazioni dei bambini con ADHD su un compito di Stroop emotivo, ha trovato che i bambini con ADHD avevano un tempo più difficile inibire le risposte ai volti arrabbiati e frustrati (Yarmolovsky et al., 2016).
Inoltre, in una meta-analisi sulla regolazione delle emozioni nell’ADHD eseguita da Graziano e Garcia (2016), hanno trovato quattro domini di interesse, ovvero: riconoscimento (cioè, capacità di elaborare e dedurre le emozioni degli altri così come il proprio sé), la reattività (cioè la soglia, l’intensità e la durata del proprio arousal affettivo), la regolazione (cioè, rispondere efficacemente alla reattività emotiva in modo flessibile che facilita il funzionamento adattivo), e l’empatia (cioè, la capacità di sperimentare lo stato affettivo di un altro e/o esprimere preoccupazione per la posizione di un altro). I risultati mostrano che mentre tutti e quattro i domini sembrano essere atipici nella popolazione ADHD, le carenze più notevoli si verificano nei domini di reazione e regolazione. Questo significa che (soprattutto) i bambini con ADHD tendono a reagire più rapidamente, più intensamente e per un periodo di tempo più lungo a situazioni avverse, e sono meno in grado di regolare queste emozioni anche quando tentano di farlo.
ADHD, selezione delle emozioni e inibizione e BA 9 laterale
Ci sono diversi studi che mostrano atipicità specifiche nell’attivazione BA 9 laterale in partecipanti ADHD che svolgono compiti che richiedono la regolazione delle emozioni. Per esempio, in uno studio di Passarotti et al. (2010a), bambini con disturbo bipolare, bambini con ADHD e un gruppo di controlli di pari età hanno eseguito un compito Stroop di valenza emotiva. In questo compito, sono state presentate parole positive, neutre e negative, e i partecipanti dovevano rispondere in base al colore della parola ignorandone il significato. I risultati mostrano che il gruppo bipolare aveva difficoltà a ignorare le parole positive, mentre il gruppo ADHD aveva difficoltà a ignorare le parole negative. È interessante notare che l’attivazione laterale BA 9 era più pronunciata per la condizione di compito eseguito con successo, cioè per le parole negative nel gruppo bipolare, e per le parole positive nel gruppo ADHD. Inoltre, in un compito emotivo basato su Stroop, Hwang et al. (2015), hanno chiesto ai bambini con e senza ADHD di eseguire un compito numerico Stroop dopo essere stato mostrato immagini che erano positive, neutre o negative. Mentre i dati comportamentali non hanno mostrato differenze significative tra le diverse condizioni emotive, hanno trovato che i bambini con ADHD erano meno in grado di reclutare il BA 9 laterale durante l’esecuzione del compito, un tratto che era anche correlato con la gravità dei sintomi come è stato misurato dalla scala dei genitori di Conner.
Motore
Selezione e inibizione motoria e BA 8 laterale
Ci sono molte prove che l’inibizione delle azioni motorie è supportata da reti neurali nel BA 8 laterale. La misura più comune dell’inibizione motoria è lo Stop Signal Task (SST) in cui i partecipanti devono premere un tasto quando viene presentato uno stimolo, ma si astengono dall’eseguire la pressione del tasto se lo stimolo è seguito a breve da un segnale. Gli studi dimostrano che l’inibizione della pressione del tasto comporta l’attivazione nella BA 8 laterale (Matthews et al., 2005; Smith et al., 2013; Hughes et al., 2014). La BA 8 laterale ha anche dimostrato di essere specificamente attiva quando la selezione di una risposta motoria richiedeva attenzione o implicava un conflitto. Per esempio, Enriquez-Geppert et al. (2013), hanno chiesto ai partecipanti di premere un tasto quando uno stimolo è apparso sullo schermo, tuttavia, il tasto di risposta è cambiato in alcune delle prove. Gli sperimentatori hanno trovato che il BA 8 laterale si attivava specificamente nelle prove in cui il tasto di risposta era diverso. Allo stesso modo, Lenartowicz et al. (2011), hanno utilizzato un compito go/no-go in cui dopo alcune sessioni di pratica, gli stimoli sono cambiati in modo che il precedente segnale go è diventato un segnale no-go (richiedendo più inibizione). Hanno trovato BA 8 laterale per essere specificamente attivato su quelle prove. In uno studio simile Albares et al. (2014), hanno usato una versione di un compito go/no-go in cui un primo è stato presentato prima degli stimoli go e no-go. Tuttavia, mentre una luce verde indicava che sarebbe seguito uno stimolo go, una luce rossa significava che poteva verificarsi una qualsiasi delle tre condizioni, uno stimolo go, uno stimolo no-go o nessuno stimolo. I ricercatori hanno ragionato sul fatto che la condizione di luce rossa comporta una pianificazione motoria più complessa rispetto alla luce verde, e hanno scoperto che il BA 8 laterale era effettivamente più attivo durante quella condizione.
ADHD e selezione e inibizione motoria
C’è una forte evidenza di deficit di selezione e inibizione motoria nei bambini con ADHD. Molti studi fino ad oggi hanno dimostrato che bambini, adolescenti e adulti con ADHD mostrano prestazioni più scarse su diverse versioni del compito del segnale di stop (ad esempio, Rubia et al., 1998; Lee et al., 2016; Bialystok et al., 2017; Dekkers et al., 2017), e che quando i bambini con ADHD sono adeguatamente medicati e motivati utilizzando un rinforzo efficace, queste differenze possono scomparire (Rosch et al., 2016). Inoltre, in uno studio molto ampio di Crosbie et al. (2013), i sintomi dell’ADHD sono stati misurati in oltre 16.000 bambini del pubblico generale che hanno eseguito la SST. I risultati hanno mostrato che gli individui con maggiori punteggi di tratti ADHD avevano una peggiore inibizione della risposta, una latenza di risposta più lenta e una maggiore variabilità, e anche che questa tendenza era altamente ereditabile. Allo stesso modo, Alderson et al. (2007), hanno eseguito una meta-analisi concentrandosi sui bambini ADHD e la SST e hanno trovato maggiori tempi medi di risposta, maggiore variabilità di risposta e maggiore tempo di risposta del segnale di arresto nel gruppo ADHD.
ADHD, selezione e inibizione motoria e BA 8 laterale
C’è anche una notevole evidenza che mostra atipicità specifiche nell’attivazione BA 8 laterale in partecipanti ADHD che eseguono compiti che richiedono selezione e inibizione motoria. Per esempio, due distinte meta-analisi hanno esplorato studi fMRI che valutano l’inibizione motoria nei bambini e negli adulti con ADHD (Hart et al., 2013; Rubia et al., 2013). Entrambe le meta-analisi hanno trovato una minore attivazione nella BA 8 laterale da correlare alle misure di inibizione motoria, e lo studio di Rubia et al. (2013), ha anche trovato il metilfenidato per aumentare l’attivazione nella BA 8 laterale che era correlata a migliori prestazioni su compiti motori. Inoltre, gli studi che hanno impiegato diverse versioni della SST e il compito go/no-go trovato meno attivazione nel BA 8 laterale nel gruppo ADHD durante una semplice condizione di no-go (Passarotti et al., 2010b; Mulligan et al., 2011), momenti prima di un errore si è verificato (Spinelli et al., 2011), così come durante una condizione di libera scelta in cui i partecipanti hanno scelto se premere un pulsante o meno quando uno stimolo è stato presentato (Karch et al, 2010).
Sommario
Il presente lavoro presenta le prove che la parte laterale della corteccia prefrontale è coinvolta nell’inibizione, selezione e manipolazione delle informazioni motorie, emotive, di memoria e percettivo-sensoriali, una funzione che è utile nella vita quotidiana attraverso la capacità di ignorare la distrazione, selezionare e recuperare informazioni specifiche dai nostri ricordi, identificare e regolare il nostro stato emotivo o pianificare una risposta motoria adeguata alla situazione. Utilizza questo modello per classificare le sfide incontrate dalle persone con ADHD, collegando diversi tipi di difficoltà nella selezione/inibizione a diverse sottoaree della corteccia prefrontale laterale.
Contributi dell’autore
L’autore conferma di essere l’unico contributore di questo lavoro e lo ha approvato per la pubblicazione.
Dichiarazione di conflitto di interessi
L’autore dichiara che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Albares, M., Lio, G., Criaud, M., Anton, J. L., Desmurget, M., e Boulinguez, P. (2014). La corteccia frontale mediale dorsale media l’inibizione automatica del motore in contesti incerti: prove da studi combinati fMRI ed EEG. Hum. Cervello Mapp. 35, 5517-5531. doi: 10.1002/hbm.22567
PubMed Abstract | CrossRef Full Text | Google Scholar
Alderson, R. M., Rapport, M. D., e Kofler, M. J. (2007). Disturbo da deficit di attenzione/iperattività e inibizione comportamentale: una revisione meta-analitica del paradigma del segnale di stop. J. Abnorm. Bambino Psychol. 35, 745-758. doi: 10.1007/s10802-007-9131-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Barkley, R. A. (1997). Inibizione comportamentale, attenzione sostenuta e funzioni esecutive: costruire una teoria unificante dell’ADHD. Psychol. Bull. 121:65. doi: 10.1037/0033-2909.121.1.65
PubMed Abstract | CrossRef Full Text | Google Scholar
Beckmann, M., Johansen-Berg, H., e Rushworth, M. F. (2009). Connettività-based parcellizzazione della corteccia cingolata umana e la sua relazione alla specializzazione funzionale. J. Neurosci. 29, 1175-1190. doi: 10.1523/JNEUROSCI.3328-08.2009
PubMed Abstract | CrossRef Full Text | Google Scholar
Bialystok, E., Hawrylewicz, K., Wiseheart, M., and Toplak, M. (2017). Interazione tra bilinguismo e disturbo da deficit di attenzione/iperattività nei giovani adulti. Bilinguismo 20, 588-601. doi: 10.1017/S1366728915000887
PubMed Abstract | CrossRef Full Text | Google Scholar
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., et al. (2014). Riappropriazione cognitiva dell’emozione: una meta-analisi di studi di neuroimaging umano. Cereb. Cortex 24, 2981-2990. doi: 10.1093/cercor/bht154
PubMed Abstract | CrossRef Full Text | Google Scholar
Castel, A. D., Lee, S. S., Humphreys, K. L., e Moore, A. N. (2011). Capacità di memoria, controllo selettivo, e valore-diretto ricordare nei bambini con e senza deficit di attenzione / iperattività (ADHD). Neuropsicologia 25:15. doi: 10.1037/a0020298
PubMed Abstract | CrossRef Full Text | Google Scholar
Cohen, M. S., Rissman, J., Suthana, N. A., Castel, A. D., e Knowlton, B. J. (2014). La modulazione basata sul valore della codifica della memoria coinvolge l’impegno strategico delle regioni di elaborazione semantica fronto-temporale. Cogn. Affect. Behav. Neurosci. 14, 578-592. doi: 10.3758/s13415-014-0275-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crosbie, J., Arnold, P., Paterson, A., Swanson, J., Dupuis, A., Li, X., et al. Inibizione della risposta e tratti ADHD: correlati ed ereditabilità in un campione di comunità. J. Abnorm. Child Psychol. 41, 497-507. doi: 10.1007/s10802-012-9693-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Dekkers, T. J., van Rentergem, J. A. A., Koole, A., van den Wildenberg, W. P., Popma, A., Bexkens, A., et al. (2017). Time-on-task effetti nei bambini con e senza ADHD: esaurimento delle risorse esecutive o esaurimento della motivazione? Eur. Child Adolesc. Psychiatry. 26, 1-11. doi: 10.1007/s00787-017-1006-y
CrossRef Full Text | Google Scholar
Depue, B. E., Burgess, G. C., Willcutt, E. G., Ruzic, L., and Banich, M. T. (2010). Controllo inibitorio del recupero della memoria e l’elaborazione motoria associata alla corteccia prefrontale laterale destra: prove da deficit in individui con ADHD. Neuropsicologia. 48, 3909-3917. doi: 10.1016/j.neuropsychologia.2010.09.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Enriquez-Geppert, S., Eichele, T., Specht, K., Kugel, H., Pantev, C., and Huster, R. J. (2013). Parcellazione funzionale delle cortecce frontali inferiori e midcingulate in un paradigma flanker-stop-cambio. Hum. Cervello Mapp. 34, 1501-1514. doi: 10.1002/hbm.22002
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., e Lavie, N. (2016). Stabilire il tratto di attenzione-distrazione. Psychol. Sci. 27, 203-212. doi: 10.1177/0956797615617761
PubMed Abstract | CrossRef Full Text | Google Scholar
Forster, S., Robertson, D. J., Jennings, A., Asherson, P., e Lavie, N. (2014). Tappare il deficit di attenzione: carico percettivo contatori maggiore distrazione in ADHD. Neuropsicologia 28:91. doi: 10.1037/neu0000020
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank, D. W., Dewitt, M., Hudgens-Haney, M., Schaeffer, D. J., Ball, B. H., Schwarz, N. F., et al. (2014). Regolazione delle emozioni: meta-analisi quantitativa di attivazione e disattivazione funzionale. Neurosci. Biobehav. Rev. 45, 202-211. doi: 10.1016/j.neubiorev.2014.06.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Gilbert, S. J., Spengler, S., Simons, J. S., Steele, J. D., Lawrie, S. M., Frith, C. D., et al. Specializzazione funzionale all’interno della corteccia prefrontale rostrale (area 10): una meta-analisi. J. Cogn. Neurosci. 18, 932-948. doi: 10.1162/jocn.2006.18.6.932
PubMed Abstract | CrossRef Full Text | Google Scholar
Graziano, P. A., and Garcia, A. (2016). Disturbo da deficit di attenzione e iperattività e disregolazione delle emozioni dei bambini: una meta-analisi. Clin. Psychol. Rev. 46, 106-123. doi: 10.1016/j.cpr.2016.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Hallam, G. P., Webb, T. L., Sheeran, P., Miles, E., Niven, K., Wilkinson, I. D., et al. (2014). I correlati neurali della regolazione delle emozioni di un’altra persona: uno studio fMRI esplorativo. Front. Hum. Neurosci. 8:376. doi: 10.3389/fnhum.2014.00376
PubMed Abstract | CrossRef Full Text | Google Scholar
Hart, H., Radua, J., Nakao, T., Mataix-Cols, D., e Rubia, K. (2013). Meta-analisi di studi di risonanza magnetica funzionale dell’inibizione e dell’attenzione nel disturbo da deficit di attenzione/iperattività: esplorando gli effetti compito-specifici, farmaci stimolanti ed età. JAMA Psychiatry. 70, 185-198. doi: 10.1001/jamapsychiatry.2013.277
PubMed Abstract | CrossRef Full Text | Google Scholar
Howard, J. D., and Kahnt, T. (2017). Identità-specifiche rappresentazioni di ricompensa nella corteccia orbitofrontale sono modulati da svalutazione selettiva. J. Neurosci. 37, 3473-3416. doi: 10.1523/JNEUROSCI.3473-16.2017
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, M. E., Budd, T. W., Fulham, W. R., Lancaster, S., Woods, W., Rossell, S. L., et al. (2014). Attivazione cerebrale sostenuta che supporta le prestazioni del compito di stop-segnale. Eur. J. Neurosci. 39, 1363-1369. doi: 10.1111/ejn.12497
PubMed Abstract | CrossRef Full Text | Google Scholar
Hwang, S., White, S. F., Nolan, Z. T., Craig Williams, W., Sinclair, S., and Blair, R. J. (2015). Controllo esecutivo dell’attenzione e risposta emotiva nel disturbo da deficit di attenzione/iperattività-Uno studio di risonanza magnetica funzionale. Neuroimage Clin. 9, 545-554. doi: 10.1016/j.nicl.2015.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Ikeda, Y., Okuzumi, H., e Kokubun, M. (2013). Stroop / reverse-Stroop interferenza nello sviluppo tipico e la sua relazione con i sintomi di ADHD. Res. Dev. Disabil. 34, 2391-2398. doi: 10.1016/j.ridd.2013.04.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Karch, S., Thalmeier, T., Lutz, J., Cerovecki, A., Opgen-Rhein, M., Hock, B., et al. Correlati neurali (ERP/fMRI) della selezione volontaria in pazienti adulti con ADHD. Eur. Arch. Psychiatry Clin. Neurosci. 260, 427-440. doi: 10.1007/s00406-009-0089-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaufman, D. A., Keith, C. M., and Perlstein, W. M. (2016). Corteccia orbitofrontale e l’elaborazione precoce della novità visiva nell’invecchiamento sano. Front. Aging Neurosci. 8:101. doi: 10.3389/fnagi.2016.00101
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2011). Attività neurale che predice la memoria successiva e dimenticare: una meta-analisi di 74 studi fMRI. Neuroimage 54, 2446-2461. doi: 10.1016/j.neuroimage.2010.09.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, H. (2013). Attività neurale differenziale nel riconoscimento di eventi vecchi rispetto a quelli nuovi: una stima di likelihood activation meta-analisi. Hum. Brain Mapp. 34, 814-836. doi: 10.1002/hbm.21474
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. Y., Wu, T. F., Tsai, J. D., and Yang, E. L. (2016). Applicabilità del compito di stop-segnale per i bambini in età prescolare con ADHD. Percept. Mot. Skills. 123, 162-174. doi: 10.1177/0031512516660715
PubMed Abstract | CrossRef Full Text | Google Scholar
Lenartowicz, A., Verbruggen, F., Logan, G. D., e Poldrack, R. A. (2011). Inibizione attivazione correlata nel giro frontale inferiore destro in assenza di spunti inibitori. J. Cogn. Neurosci. 23, 3388-3399. doi: 10.1162/jocn_a_00031
PubMed Abstract | CrossRef Full Text | Google Scholar
Matthews, S. C., Simmons, A. N., Arce, E., e Paulus, M. P. (2005). Dissociazione dell’inibizione dall’elaborazione degli errori utilizzando un compito inibitorio parametrico durante la risonanza magnetica funzionale. Neuroreport. 16, 755-760. doi: 10.1097/00001756-200505120-00020
PubMed Abstract | CrossRef Full Text | Google Scholar
Mulligan, R. C., Knopik, V. S., Sweet, L. H., Fischer, M., Seidenberg, M., e Rao, S. M. (2011). Correlati neurali del controllo inibitorio nel disturbo da deficit di attenzione/iperattività dell’adulto: prove dal campione longitudinale di Milwaukee. Psychiatry Res. 194, 119-129. doi: 10.1016/j.pscychresns.2011.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Nigg, J. T. (1999). Il deficit di inibizione della risposta ADHD come misurato dal compito di arresto: replica con DSM-IV tipo combinato, estensione e qualificazione. J. Abnorm. Child Psychol. 27, 393-402. doi: 10.1023/A:1021980002473
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010a). Correlati neurali di inibizione della risposta nel disturbo bipolare pediatrico e disturbo da deficit di attenzione e iperattività. Psichiatria Res. Neuroimaging. 181, 36-43. doi: 10.1016/j.pscychresns.2009.07.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Passarotti, A. M., Sweeney, J. A., and Pavuluri, M. N. (2010b). Impegno differenziale dei sistemi neurali cognitivi e affettivi nel disturbo bipolare pediatrico e nel disturbo da deficit di attenzione e iperattività. J. Int. Neuropsychol. Soc. 16, 106-117. doi: 10.1017/S1355617709991019
PubMed Abstract | CrossRef Full Text | Google Scholar
Pelletier, M. F., Hodgetts, H. M., Lafleur, M. F., Vincent, A., and Tremblay, S. (2016). Vulnerabilità all’effetto sonoro irrilevante nell’adulto ADHD. J. Atten. Disord. 20, 306-316. doi: 10.1177/1087054713492563
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollak, Y., Kahana-Vax, G., e Hoofien, D. (2007). Processi di recupero in adulti con ADHD: uno studio RAVLT. Dev. Neuropsychol. 33, 62-73. doi: 10.1080/87565640701729789
PubMed Abstract | CrossRef Full Text | Google Scholar
Pretus, C., Picado, M., Ramos-Quiroga, A., Carmona, S., Richarte, V., Fauquet, J., et al. (2016). Presenza di distrattore migliora le prestazioni di stima del tempo in un campione adulto ADHD. J. Atten. Disord. doi: 10.1177/1087054716648776. .
PubMed Abstract | CrossRef Full Text | Google Scholar
Price, J. L. (2007). Definizione della corteccia orbitale in relazione alle connessioni specifiche con strutture limbiche e viscerali e altre regioni corticali. Annals N.Y. Acad. Sci. 1121, 54-71. doi: 10.1196/annals.1401.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Rabinak, C. A., MacNamara, A., Kennedy, A. E., Angstadt, M., Stein, M. B., Liberzon, I., et al. (2014). Impegno prefrontale focale e aberrante durante la regolazione delle emozioni nei veterani con disturbo da stress post-traumatico. Depressione. Ansia. 31, 851-861. doi: 10.1002/da.22243
PubMed Abstract | CrossRef Full Text | Google Scholar
Reimer, B., Mehler, B., D’Ambrosio, L. A., e Fried, R. (2010). L’impatto delle distrazioni sui conducenti giovani adulti con disturbo da deficit di attenzione e iperattività (ADHD). Accid. Anal. Prev. 42, 842-851. doi: 10.1016/j.aap.2009.06.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosch, K. S., Fosco, W. D., Pelham, W. E. Jr., Waxmonsky, J. G., Bubnik, M. G., e Hawk, L. W. Jr. (2016). Il rinforzo e i farmaci stimolanti migliorano la carente inibizione della risposta nei bambini con disturbo da deficit di attenzione/iperattività. J. Abnorm. Child Psychol. 44, 309-321. doi: 10.1007/s10802-015-0031-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Lim, L., Ecker, C., Halari, R., Giampietro, V., Simmons, A., et al. Effetti di età e di genere sulle reti neurali di inibizione della risposta motoria: dall’adolescenza alla metà dell’età adulta. Neuroimage. 83, 690-703. doi: 10.1016/j.neuroimage.2013.06.078
PubMed Abstract | CrossRef Full Text | Google Scholar
Rubia, K., Oosterlaan, J., Sergeant, J. A., Brandeis, D., e Leeuwen, T. V. (1998). Disfunzione inibitoria in ragazzi iperattivi. Comportamento. Brain Res. 94, 25-32. doi: 10.1016/S0166-4328(97)00166-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Shalom, D. B. (2009). La corteccia prefrontale mediale e l’integrazione nell’autismo. Neuroscientist. 15, 589-598. doi: 10.1177/1073858409336371
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, J. L., Jamadar, S., Provost, A. L., e Michie, P. T. (2013). Inibizione motoria e non-motoria nel compito Go/NoGo: uno studio ERP e fMRI. Int. J. Psychophysiol. 87, 244-253. doi: 10.1016/j.ijpsycho.2012.07.185
PubMed Abstract | CrossRef Full Text | Google Scholar
Soliman, A. M., and Elfar, R. M. (2017). Memoria falsa in adulti con ADHD: un confronto tra sottotipi e controlli normali. J. Attent. Disord. 21, 986-996. doi: 10.1177/1087054714556814
PubMed Abstract | CrossRef Full Text | Google Scholar
Sørensen, L., Plessen, K. J., Adolfsdottir, S., e Lundervold, A. J. (2014). La specificità del punteggio di interferenza Stroop di errori all’ADHD nei ragazzi. Bambino Neuropsychol. 20, 677-691. doi: 10.1080/09297049.2013.855716
PubMed Abstract | CrossRef Full Text | Google Scholar
Soto, D., Humphreys, G. W., e Rotshtein, P. (2007). Dissociare i meccanismi neurali di guida basata sulla memoria della selezione visiva. Proc. Natl. Acad. Sci. U.S.A. 104, 17186-17191. doi: 10.1073/pnas.0703706104
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinelli, S., Joel, S., Nelson, T. E., Vasa, R. A., Pekar, J. J., e Mostofsky, S. H. (2011). Diversi modelli neurali sono associati con le prove che precedono gli errori inibitori nei bambini con e senza disturbo da deficit di attenzione/iperattività. J. Am. Acad. Child Adolesc. Psychiatry. 50, 705-715. doi: 10.1016/j.jaac.2011.03.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Tsujimoto, S., Yasumura, A., Yamashita, Y., Torii, M., Kaga, M., e Inagaki, M. (2013). Aumento dell’ossigenazione prefrontale legata alla memoria di lavoro resistente ai distrattori nei bambini con disturbo da deficit di attenzione/iperattività (ADHD). Psichiatria infantile Hum. Dev. 44, 678-688. doi: 10.1007/s10578-013-0361-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Walcott, C. M., and Landau, S. (2004). La relazione tra disinibizione e regolazione delle emozioni nei ragazzi con disturbo da deficit di attenzione e iperattività. J. Clin. Child Adolesc. Psychol. 33, 772-782. doi: 10.1207/s15374424jccp3304_12
PubMed Abstract | CrossRef Full Text | Google Scholar
Xiong, K., Zhang, Y., Qiu, M., Zhang, J., Sang, L., Wang, L., et al. Regolazione delle emozioni negative in pazienti con disturbo da stress post-traumatico. PLoS ONE 8:e81957. doi: 10.1371/journal.pone.0081957
PubMed Abstract | CrossRef Full Text | Google Scholar
Yarmolovsky, J., Szwarc, T., Schwartz, M., Tirosh, E., e Geva, R. (2016). Controllo esecutivo caldo e risposta a uno stimolante in uno studio randomizzato in doppio cieco in bambini con ADHD. Eur. Arch. Psychiatry Clin. Neurosci. 267, 73-82. doi: 10.1007/s00406-016-0683-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Yasumura, A., Kokubo, N., Yamamoto, H., Yasumura, Y., Nakagawa, E., Kaga, M., et al. (2014). Valutazione neurocomportamentale ed emodinamica di Stroop e interferenza Stroop inversa nei bambini con disturbo da deficit di attenzione/iperattività. Cervello Dev. 36, 97-106. doi: 10.1016/j.braindev.2013.01.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Zsuga, J., Biro, K., Tajti, G., Szilasi, M. E., Papp, C., Juhasz, B., et al. (2016). Uso ‘proattivo’ della congruenza cue-contesto per la costruzione della funzione di ricompensa dell’apprendimento di rinforzo. BMC Neurosci. 17:70. doi: 10.1186/s12868-016-0302-7
PubMed Abstract | CrossRef Full Text | Google Scholar