The Brazil Nut Industry — Past, Present, and Future
Scott A. Mori
The New York Botanical Garden
この記事の転載は Sustainable Harvest and Marketing of Rain Forest Productsの許可を得て行っています。 Plotkin, M. and L. Famolare (eds.). Copyright Island Press, 1992. 発行:Island Press, Washington, D.C. & Covelo, California.発行:Island Press, Washington, D.C. & Covelo, California. 本書のご注文は、1-800-828-1302にお電話いただくか、下記まで書面にてお願いいたします。 Island Press, Box 7, Covelo, CA 95428
はじめに|自然史|フェノロジー|受粉生物学|散布生物学|ブラジルナッツ収穫|
ブラジルナッツプランテーション|ブラジルナッツ生産の将来|参考文献
はじめに
アマゾンの最も重要な経済的植物の1つは、ブラジルナッツ(Bertholletia excelsa、レシチダ科)です。 この種の食用種子は、Hevea brasiliensisのラテックスとともに、しばしばアマゾンの抽出埋蔵量の最も重要な産物として挙げられています。 ブラジルのナッツは主に雨季に採取され、ゴムは主に乾季に採取される。 この2つの林産物の組み合わせは、採収活動で生活する人々にとって年間を通じての収入となります。 ブラジルナッツやゴムの採取は、アマゾンの森林の生態系に比較的影響を及ぼしにくい。 そのため、生物多様性の保全と、これらやその他の非木材製品の開発は両立するとよく言われます。 しかし、非木材林産物を採取する人々は、ほとんどの場合、焼畑農業、木材採取、鉱業、狩猟などの他の活動に関わっています。 その結果、頻繁に利用される採掘保護区では、アマゾンの森林に見られる膨大な生物多様性の一部しか保護されていないのが普通です。 したがって、アマゾンの保全計画には、アマゾンのすべての生態系を過度の経済的搾取から保護する大規模な保護区を含める必要があります。
この章では、ブラジルナッツの自然史、収穫の価値、プランテーション栽培の可能性、そしてブラジルナッツ産業の将来について概説します。 ブラジルナッツはその経済的重要性から、その生物学や農学に関する多くの研究の対象となってきた。 その中でも、ブラジルのパラー州ベレンにあるEMBPRAPA(Empresa Brasileira de Pesquisas Agropecuária)のCPATU(Centro de Pesquisa Agropecuária do Trópico Umido)が最も多くの研究をしている。 ブラジルナッツに関する文献は259タイトルあり(Vaz Pereira and Lima Costa, 1981)、ブラジルナッツの生物学および農学に関する最近のまとめはMori and Prance (1990b)に掲載されている。
分類と分布
Bertholletia excelsaは汎熱帯樹科(Lecythidaceae)に属し、新熱帯には約200種がメキシコ南部からブラジル南部まで分布する(Mori and Prance, 1990a; Prance and Mori, 1979)。 ブラジルナッツはBertholletia属の1種であり、その定義が明確である。 果実の大きさや形、果実あたりの種子数にはかなりの変異があるが、Bertholletia属を複数種認める正当な理由はない。
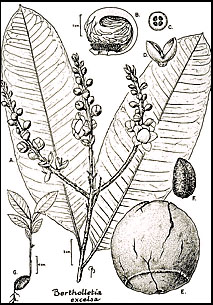 B. excelsaの近縁種は,俗にjarana groupと呼ばれるLecythisの種の中にある(Mori and Prance, 1990b)。 Lecythis pisonisとその近縁種のL. minorとL. ollariaもこの仲間で,種子を食用とする他の種である。 しかし、後者2種は、種子中の過剰なセレン蓄積により、時に毛髪や爪の脱毛を引き起こす (Dickson, 1969; Kerdel-Vegas, 1966)。 しかし、BertholletiaとLecythisの違いは非常に大きく、交雑によって一方の属から他方の属に遺伝物質を導入することはほとんど期待できない。 したがって、ブラジルナッツ生産の改良のための生殖質は、他の属の近縁種からではなく、B. excelsaの中に見られる変異から得なければならないだろう。
B. excelsaの近縁種は,俗にjarana groupと呼ばれるLecythisの種の中にある(Mori and Prance, 1990b)。 Lecythis pisonisとその近縁種のL. minorとL. ollariaもこの仲間で,種子を食用とする他の種である。 しかし、後者2種は、種子中の過剰なセレン蓄積により、時に毛髪や爪の脱毛を引き起こす (Dickson, 1969; Kerdel-Vegas, 1966)。 しかし、BertholletiaとLecythisの違いは非常に大きく、交雑によって一方の属から他方の属に遺伝物質を導入することはほとんど期待できない。 したがって、ブラジルナッツ生産の改良のための生殖質は、他の属の近縁種からではなく、B. excelsaの中に見られる変異から得なければならないだろう。
Bertholletia excelsaはアマゾンの植物で、ギアナ、コロンビア、ベネズエラ、ペルー、ボリビア、ブラジルの非洪水林(テラファーマ)を好む。 ブラジルナッツが生育する気候条件は、de Almeida (1963), Diniz and Bastos (1974), および Mori and Prance (1990b) にまとめられている。
ブラジルナッツは自生地以外の熱帯植物園で栽培されており、マレーシアのクアラルンプール (Müller, 1981) とアフリカのガーナ (D.K. Abbiw, pers. comm.) に小規模なプランテーションが確立されている。
ペルーではmanchales (Sánchez, 1973)、ブラジルではcastanhais (Dias, 1959) と呼ばれる50~100個体の樹林が存在し、この樹林の中にBertholletia excelsaの木がある。 アマゾン全域で、1ヘクタールあたりのブラジルナッツの木の密度はかなり異なっている。 ブラジル東部のブラジルナッツ生産に関する研究では、Miller(1990)は1ヘクタールあたり9~26本の繁殖木を発見したが、BeckerとMori(未発表データ)はブラジル中央部の100ヘクタールの区画で10cm dbh以上の木を1本だけ発見している。
ブラジルナッツの木はギャップ依存性であるという証拠がいくつかある(Mori and Prance, 1990b)。 さらに、ブラジルナッツの木立は、コロンビア以前のインディアンに起源があると示唆する著者もいる(Miller, 1990; Mori and Prance, 1990b; Müller et al.) ブラジルナッツの自然林の管理を可能にするためには、種子からの繁殖個体の発生について理解することが必要である。
Bertholletia excelsaの開花は、乾季から雨季にかけて行われる。 実際、ブラジルナッツは3~5ヶ月の乾季がある地域でのみ自生している(Müller, 1981)。 アマゾン東部では、開花は雨季の終わりの9月に始まり、2月まで続く。 開花のピークは10月、11月、12月である(Moritz, 1984)。
雨季の終わり頃、一般的には7月になると、ブラジルナッツの木の葉は落ち始める。 前年の花序の真下から新芽が吹き出し、その頂点に新しい花序ができる。 毎日、比較的長い期間、大量の花を咲かせる。 花は午前4時半から5時の間に開くが、葯は花が開く数時間前に蕾の中で脱皮し始める。 花弁と葯は花が開いた日の午後に落下する (Mori and Prance, 1990b) 。
果実の成熟はレシチン科の他のどの種よりも長い時間をかけて行われる。 Moritz (1984)は、結実してから果実が展開するまでに15ヶ月を要するとしている。 そのため、ブラジルナッツの果実は、雨季の1月から2月に多く落下する。 自然条件下では、種子の発芽に12~18ヶ月かかる(Müller, 1981)。
 ブラジルナッツの花は接合型で、片側が伸びてアーチ状になった被子植物が卵巣の頂部にぴったりとくっ付いています。 また、花弁は雌しべに密着している(図27-1)。 そのため、花に入ることができるのは、アンドロメダ頭巾をこじ開けるだけの力を持った大きな体のハチだけであり、コイル状のアンドロメダ頭巾の頂点に生じる蜜と思われる受粉の報酬を手に入れることができる。 Bombus属,Centris属,Epicharis属,Eulaema属,Xylocopa属のハチがブラジルナッツの木を訪れるのが確認されている (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985)。 これらのハチは非社会性もしくは半社会性であり、社会性ハチ (例えば Apis, Melipona, Trigona) のように、ハチの巣を農園から農園に運ぶことで特定の作物の受粉に利用できるような、人間による操作を容易に行うことができない。
ブラジルナッツの花は接合型で、片側が伸びてアーチ状になった被子植物が卵巣の頂部にぴったりとくっ付いています。 また、花弁は雌しべに密着している(図27-1)。 そのため、花に入ることができるのは、アンドロメダ頭巾をこじ開けるだけの力を持った大きな体のハチだけであり、コイル状のアンドロメダ頭巾の頂点に生じる蜜と思われる受粉の報酬を手に入れることができる。 Bombus属,Centris属,Epicharis属,Eulaema属,Xylocopa属のハチがブラジルナッツの木を訪れるのが確認されている (Moritz, 1984; Müller et al., 1980; Nelson et al., 1985)。 これらのハチは非社会性もしくは半社会性であり、社会性ハチ (例えば Apis, Melipona, Trigona) のように、ハチの巣を農園から農園に運ぶことで特定の作物の受粉に利用できるような、人間による操作を容易に行うことができない。
 新熱帯のレシチン科では、ほとんどの場合、種子の着床に他家受粉が必要である。 したがって、レシチダスの受粉とそれに続く果実や種子の発育には、ハチ、そして、より少ない範囲ではあるがコウモリが不可欠である。 Bertholletia excelsaでは低レベルの同系交配が起こるかもしれないが,この種の種子形成のほとんどは他家受粉の結果である (Mori and Prance, 1990b)。 ブラジルナッツの自家和合性系統の開発は、管理が難しいハチ受粉媒介者による他花受粉の必要性を排除することで、この種のプランテーション栽培を容易にする。
新熱帯のレシチン科では、ほとんどの場合、種子の着床に他家受粉が必要である。 したがって、レシチダスの受粉とそれに続く果実や種子の発育には、ハチ、そして、より少ない範囲ではあるがコウモリが不可欠である。 Bertholletia excelsaでは低レベルの同系交配が起こるかもしれないが,この種の種子形成のほとんどは他家受粉の結果である (Mori and Prance, 1990b)。 ブラジルナッツの自家和合性系統の開発は、管理が難しいハチ受粉媒介者による他花受粉の必要性を排除することで、この種のプランテーション栽培を容易にする。
ブラジルナッツの自生域以外のハチも受粉に影響を与えることができる。 例えば、セイロン(Macmillan, 1935)、クアラルンプール、ガーナのブラジルナッツの木は実をつける。
 B. excelsaの果実と種子はレシチン科の中ではユニークなもので、果実と種子は、レシチン科の中でも特に優れています。 成熟すると、大きく丸い木質の果実は、中に種子を含んで地面に落ちる。 種子は骨質で,莢から取り出され,齧歯類,特にアグーチ(Dasyprocta spp.)によって散布される (Huber, 1910; Miller; 1990; E. Ortiz, pers. comm.). アグーチスとリスは,非常に木質化した周皮を効率的にかじることができる唯一の動物であると思われる。 彼らは種子の一部を食べ、残りの種子は後で使用するために貯蔵する。
B. excelsaの果実と種子はレシチン科の中ではユニークなもので、果実と種子は、レシチン科の中でも特に優れています。 成熟すると、大きく丸い木質の果実は、中に種子を含んで地面に落ちる。 種子は骨質で,莢から取り出され,齧歯類,特にアグーチ(Dasyprocta spp.)によって散布される (Huber, 1910; Miller; 1990; E. Ortiz, pers. comm.). アグーチスとリスは,非常に木質化した周皮を効率的にかじることができる唯一の動物であると思われる。 彼らは種子の一部を食べ、残りの種子は後で使用するために貯蔵する。
 ブラジルナッツは雨季の5~6ヶ月間に野生の木からほぼ完全に収穫される。 果実は0.5~2.5kgで、10~25個の種子を含んでいるが、種子に対する虫やカビの攻撃を最小限に抑え、動物によって運ばれる種子数をコントロールするために、落下後すぐに集められる (Mori and Prance, 1990b)。 Miller (1990)によれば,1本の木にできる莢の数は63から216個である。 ブラジルナッツの収穫方法についてのより詳細な記述は、Almeida (1963), Mori and Prance (1990b), および Souza (1963)に掲載されている。
ブラジルナッツは雨季の5~6ヶ月間に野生の木からほぼ完全に収穫される。 果実は0.5~2.5kgで、10~25個の種子を含んでいるが、種子に対する虫やカビの攻撃を最小限に抑え、動物によって運ばれる種子数をコントロールするために、落下後すぐに集められる (Mori and Prance, 1990b)。 Miller (1990)によれば,1本の木にできる莢の数は63から216個である。 ブラジルナッツの収穫方法についてのより詳細な記述は、Almeida (1963), Mori and Prance (1990b), および Souza (1963)に掲載されている。
ブラジルナッツの採集は、アマゾンの地元経済に大きな影響を与える。 しかし、アマゾンから正確なデータを得ることが困難であるため、入手可能な数字は総生産量の概算を示すにすぎない。 ブラジルの生産量は、1944年の3,557トンから1970年の104,487トンまで幅があります。 1980 年以降、年間生産量は約 40,000 トンで推移している(Mori and Prance, 1990b)。 かつて、ペルーのプエルト・マルドナード(Sánchez, 1973)やブラジルのマラバ(Dias, 1959)など、アマゾンの多くの町の福祉は、ブラジルナッツの生産に大きく依存していた。 1986 年、マナウスから輸出された殻付きおよび殻なしのブラジルナッツ種子の総額は、5,773,228 ドルであった。 (Mori and Prance, 1990b). 種子の多くは、イギリス、フランス、アメリカ、ドイツに送られる。
ミラー(1990)による計算では、ブラジルナッツスタンドの一次価値(採集者に支払われるお金)は、1ヘクタールあたり97ドルと推定されている。 この価値には、スタンドに残された種子を考慮し、任意の25%割引が含まれています。 二次的価値とは、米国を拠点とする輸入会社が殻を剥いたナッツを輸出する際に受け取る金銭で、1ヘクタール当たり175.56ドルと推定された。 また、フロリダのスーパーマーケットで売られている殻付きナッツ1袋の値段は、1ヘクタール当たり1059.44ドルである。 殻付きナッツや加工ナッツの方がはるかに価値が高い。 カルチュラル・サバイバル社のように、殻むきと加工の一部を採集者のコミュニティで行う取り組みは、採収を維持するためのさらなるインセンティブになる。 10年間にわたり、ブラジルナッツの生産に森林を利用することは、木材の採取や牧草地として森林を伐採するよりも収益性が高いようです(Miller, 1990)。
ブラジルナッツのプランテーション
大規模プランテーションでのブラジルナッツの栽培方法は、ブラジル、ベレンのCPATUのMüllerとその仲間によって開発されました。 彼らの研究の英文概要はMori and Prance (1990b)に記載されている。 前述したように、ブラジルナッツの生産の大部分はまだ野生樹から採取されている。 しかし、amazonの各地ではプランテーションが開発されている。 1990年1月、私はブラジル・アマゾナス州のマナウス/イタコアティアラ街道の215km地点にあるブラジルナッツのプランテーション、Fazenda Aruanãを訪ねた。 ここは、1980年に一部ブラジルナッツ農園に転換した12,000ヘクタールの元牧場である。 私が訪れた時には、3,341ヘクタールの土地に318,660本のブラジルナッツが植えられていた。 また、近隣のFazenda Aruanãの森林には、未知数の個体が生育していた。
Fazenda Aruanãの当初の目的は、20×20メートルのグリッドにブラジルナッツの木を植え、その木の間に牛を放牧することでした。 当初、牛は木の葉を食べなかったが、後に食べるようになった。 その結果、10m×10mに間隔を狭め、牛の頭数も300頭に減らした。 アルアナン植林地の木は、ブラジルナッツの大果実と種子で知られるアブファリ・アマゾナス地方の高収量クローンを接ぎ木したものである。 このプランテーションや他のプランテーションを設立する際に重要なことは、種子の産地である。 Moritz(1984)は、同一クローン間の受精の結果としての果実生産が少ないことを明らかにしている。 この考え方に基づき、Müller(1981)は接ぎ木用の芽は少なくとも5本の異なる樹木から得ることを推奨している。 接ぎ木に使うクローンの数が少ないと、果実の収量が多いものを選ぶと、将来の病気や害虫の攻撃に対して農園の抵抗力が弱くなるという危険もある。
ファゼンダ・アルアナンでは、種子から根株を栽培している。 種子は湿った砂の中で軟化させ、その後、種皮を縁に沿って開いて胚を取り出すことで発芽させる。 殺菌剤で処理された胚は約20日で発芽し、ビニール袋やカップで苗を育てる。 畑に移植する際には、苗を植える穴に約200gのリン酸を加える。 根株は、幼木が畑によく根付く1年半から2年ぐらいで接ぎ木ができるようになる。 穂木は8~10日前に枝から葉を取り除くと、より良いものが得られる。 こうすることで、接ぎ木に適した元気な芽が形成される。 接ぎ木をしてから30日後に、根株を接ぎ木の遠位に輪生させる。 こうすることで、穂木は徐々に根株の枝を伸ばしていく。 6年後、実がなり始める。 しかし、私が訪問した時には、それまで実った果実はすべて、根株を増やすか、苗木を生産し、他の人に販売するために使用されていた。
この農園では、受粉媒介者の不足による問題はないようだ。 ビシャ・オレラナは受粉媒介者の餌となる花粉を供給するために植えられたのである。 また、プランテーションの周囲には広大な自然林があり、自然植生の風穴がプランテーションを横切っている。 風穴には、ブラジルナッツの受粉媒介者の餌となる植物が多数植えられている。 当時は樹齢が10年近くありましたが、病気の兆候や履歴はありませんでした。
ファゼンダ・アルアナン社のオーナーは、この農園に大きな期待を寄せています。 12年後の収穫量は1本あたり4.8kg、1ヘクタールあたり480ドルを見込んでいます。 18年後には1本あたり8.5キログラム、1ヘクタールあたり850ドルになると予測しています。
ブラジルナッツ生産の将来
これまでブラジルナッツは、ほとんどが野生の木から採取されていた。 近年、ブラジルナッツの生産は、森林伐採、ブラジルナッツ採集者の大都市への流出、いくつかの伝統的なブラジルナッツの木の洪水、そしておそらくブラジルナッツの木の花が咲く乾季に火災によって花粉媒介者が混乱したために減少した(Kitamura and Müller, 1984; Mori and Prance, 1990b)。 伝統的なブラジルナッツの生産方法を維持するためには、ブラジルナッツの木の密度が高い地域に非常に大規模な採収施設を設置する必要がある。
しかし、熱帯の自然保護に関心を持つ人々は、採取保護区の設立をアマゾンの生物多様性の維持と同一視しないように注意しなければならない。 なぜなら、ブラジルナッツ採集者やゴムノキ栽培者は、単にブラジルナッツを採集するだけではなく、植物や動物の多様性にマイナスの影響を与えることが多いからです。 ブラジルナッツやゴムの木のような経済的な植物があちこちに点在し、二次的な植生に過ぎなくなる可能性があります。 したがって、採掘権の保護区を設定することは、十分に計画された生物保護区の必要性を否定することにはならない。
ブラジルナッツのプランテーションが将来的に成功するかどうかは、まだ議論の余地がある。 これまで、このアマゾンの木のプランテーションが経済的に成功した例はない。 自然にはほぼ単一型の樹形で生育しない熱帯樹をプランテーションで育てる試みは、すべて失敗しているのである。 ブラジルのナットは、ゴムの木と同じように、森の中に多かれ少なかれ点在している。 ゴムの生息域内でゴムをプランテーション化しようとしたことが経済的に大失敗したことはよく知られており(Hecht and Cockburn, 1989)、アマゾンのブラジルナッツのプランテーションがアマゾンのゴムプランテーションと同じ運命にならないと信じる理由はないだろう。 今後10年間、アルアナン農園を注意深く観察することで、農園でのブラジルナッツ生産が野生木からの採集に代わる実行可能な方法かどうかを判断することができるかもしれない。 もし、プランテーションが実行可能であれば、自然保護活動家は、プランテーションが抽出保護区の維持に与える影響を評価する準備をしなければならないだろう。
最後に、アマゾンで増え続ける人口を支える経済的に実行可能な方法として、ブラジルナッツの抽出に過度の期待をかけないことが重要である。 第一に、世界の市場はブラジルナッツの生産量の増加に対応できないかもしれないし、第二に、このような低強度の土地利用は、より多くの人々が求める生活水準の向上に必要なレベルで、人間集団を支えることができないからである。 狩猟採集と焼畑農業がそれぞれ5平方キロメートルあたり1人、1平方キロメートルあたり1人しか養えないとするEwel(1991)の推定が正しいとすれば(そうでないと考える理由はない)、採掘権の保護はアマゾン諸国の人口増加をほとんど吸収することができないだろう。 抽出埋蔵量の将来と、新熱帯の生物多様性の代表サンプルを保存するための人類の能力は、熱帯の内外で人口増加と消費をうまくコントロールできるかどうかにかかっている(Erhlich and Erhlich, 1990)。
de Almeida, C.P. 1963. Castanha do Para: Sua exportação e importância na economia Amazonica. ブラジルの研究 19:1-86.
Balick, M.J. 1985. アマゾンの有用植物. 世界的に重要な資源. G.T.Prance and T.E. Lovejoy (eds.), Amazonia.にて. New York: Pergamon Press.
Dias, C.V. 1959. トカンチンス地方におけるカスターニャ産業の地理的側面. Revista Brasil. 21(4):77-91.
Dickson, J.D. 1969. Sapucaia Nuts (Lecythis elliptica)を摂取した後の髪と爪の損失に関するノート. Econ. Bot. 23:133-34.
de Diniz, T.D., A. S. Diniz, and T. X. Bastos. 1974. ブラジルのヒマワリの典型的な気候に関する知見への貢献. Bol. Técn. ipean 64:59-71.
Ewel, J. 1991. 保全と農業. Tropinet 2(1):1.
Ehrlich, A.H., and P.R. Ehrlich. 1990. Extinction: 危機に瀕した生命。 S. Head and R. Heinzman (eds.), Lessons of the Rainforest.による。 サンフランシスコ。 Sierra Club Books.
Farnsworth, N.R. 1984. 井戸は水で満たされているのに、どうして枯れているのか? Econ. Bot. 38:4-13.
ヘクト,S.,A.コックバーン. 1989. 森林の運命. New York: Verso.
Huber, J. 1910. マッタスとマデイラ・アマゾニカ. Bol. Mus. Paraense Hist. Nat. 6:91-225.
Kerdel-Vegas, F. 1966. ココ・デ・モノ(Lecythis ollaria)の脱毛および細胞毒性作用と慢性皮膚病との関連. Econ. Bot. 20:187-95.
Kitamura, P.C., and C.H. Müller. 1984. マラバの在来種カスタニウス. その保存のための基礎知識. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido(海底油田農業研究センター)。 をご覧ください。
Macmillan, H.F. 1935. 熱帯の植栽と園芸-セイロンを中心に-. London: マクミラン.
Menezes, M.A. 1990. (Reservas extrativistas: Por uma reforma agrária ecológica.) Ciência Hoje 11(64):4-6.
Miller, C. 1990. ブラジルナッツの自然史,経済植物学,および生殖質保全(Bertholletia excelsa Humb. and Bonpl). フロリダ大学修士論文.
Mori, S. A., and G.T. Prance. 1990a. レシチダス科–そのII. 接合型花をもつ新世界の属(Bertholletia, Corythophora, Couratari, Couroupita, Eschweilera, and Lecythis). Fl. ネオトープ Monogr.21(II): 1-376.
Mori, S. A., and G.T. Prance. 1990b. ブラジルナッツの分類,生態,経済植物学(Bertholletia excelsa Humb. and Bonpl.: Lecythidaceae). を紹介した。 Bot. 8:130-50.
Moritz, A. 1984. ブラジルナッツ(Bertholletia excelsa H.B.K.)の生物学的研究. EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido(海底油田農業研究センター)。 ドキュメント 29:1-82.
Müller, C. H. 1981. ブラジルナッツ、農学研究 EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido(海底油田農業研究センター)。 ドキュメント 2: 1-25.
Müller, C. H., I. A. Rodriques, A. A. Müller, and N.R.M. Müller. 1980. ブラジルナッツ 研究成果 EMBRAPA, Centro de Pesquisa Agropecuária do Trópico Umido. 雑学2:1-25。
Nelson, B. W., M. L. Absy, E. M. Barbosa, and G. T. Prance. 1985. Bertholletia excelsa H.B.K. とCouratari tenuicarpa A. C. Sm.の花客に関する観察。 (Lecythidaceae) を使用しています。 アクタ・アマゾニア 15 (1/2):225-34.
Peters, C.M., A. H. Gentry, and R.O. Mendelsohn. 1989. アマゾンの熱帯雨林の価値評価。 Nature 339:655-56。
Prance, G.T., and S.A. Mori. 1979. レシチダス科–第1部:アクティノモルフの新世界レシチダス科(Asteranthos, Gustavia, Grias, Allantoma, Cariniana). Fl. Neotrop. Monogr.21(1):1-270。
Sánchez. 1973. マドレ・デ・ディオスにおけるブラジルナッツの搾取と販売。 農林省森林・狩猟総局、報告書第20号。 ペルー・リマ
Souza A. H. 1963. カスタニャ・ド・パラー 植物学的、技術的な研究。 Ediçoes S.I.A., Estudos Técnicos 23:1-69.
Vaz Pereira, I.C., and S.L. Lima Costa. 1981. Castanha-do-Pará (Bertholletia excelsa H.B.K.) のビブリオグラフィア。 EMPRAPA, Centro de Pesquisa Agropecuária do Trópico Umido(海底油田農業研究センター)。 Belém, Pará.
Lecythidaceae Pages Home
Return to Mori Home
2002年2月設立。 問題やご質問は、[email protected] までご連絡ください。