進化と自然選択
Nature encourage no looseness, pardon no errors
– Ralph Waldo Emerson
I have called this principle, by each slight variation, if useful is preserved, by term Natural Selection.この原則を、私は、自然選択という言葉で呼びました。
– Charles Darwin, The Origin of Species
|
2010/10/10 |
|
この授業では、
- 進化論の策定に自然の中の観察結果はいかにして導かれたか?
- ダーウィンの進化論の要点は何ですか。
- 自然淘汰の過程はどのように行われるのでしょうか。
- 局所適応の根拠は?
- 自然淘汰はどのように形質の頻度に影響を与えることができるのでしょうか。
The (R)Evolution of Theory
進化論は人類史における偉大な知的革命の一つで、世界とその中での自分の位置づけについての認識を大きく変えました。 チャールズ・ダーウィンは、一貫した進化論を打ち出し、この理論を支持する多くの証拠を集めました。 ダーウィンの時代には、ほとんどの科学者が、それぞれの生物とそれぞれの適応は創造主の作品であると完全に信じていた。 リンネウスは、今日私たちが使用している生物学的分類体系を確立し、神の創造物を目録化する精神でそれを行った。
言い換えれば、生命の大樹(図1参照)を生み出す分岐プロセスの結果である生物群間の類似性と非類似性のすべてを、19世紀初期の哲学者や科学者は、全能の設計の結果と見なしたのである。
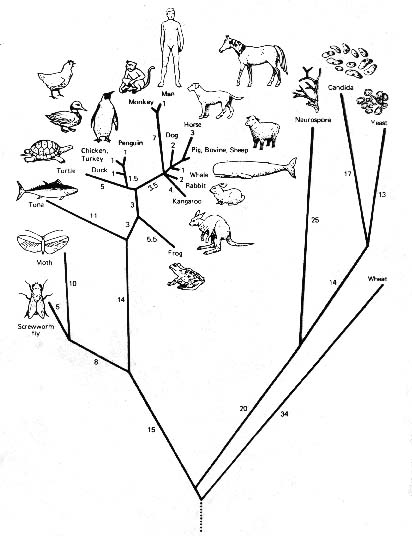
Figure 1: 示した生物のチオクロムc分子のコンピュータ解析によって構築された系統樹「生命の木」。
しかし、19世紀には、多くの自然史家が、自然界で観察されるパターンの説明として進化的変化を考え始めていたのである。 以下のような考え方は、ダーウィンの時代の知的風潮の一部であった。
- 地球が何歳か誰も知らなかったが、地質学者は地球が聖書の創造で説明されるよりかなり古いと推定し始めた。 地質学者は、地層、つまり堆積物の連続した堆積期間によって形成された層について、より深く学んでいました。 このことは、若い地層が古い地層の上に重なっているという時間的順序を示唆しています。
- 統一主義と呼ばれる概念は、地質学者チャールズ・ライエルに大きく影響されたもので、浸食や堆積物の堆積など、現在進行中で観察可能なプロセスを調査し、現在の条件とプロセスが過去への鍵であるという作業仮説に基づいて、地球の歴史を解読することを試みました。
- 18世紀から19世紀にかけては、化石の発見が相次ぎました。 当初、自然科学者は、未知の種がまだ生きている遺骸を発見したのだと考えていた。 しかし、化石の発見が進むにつれて、巨大恐竜のようなものは地球上のどこにも知られていないことが明らかになった。 さらに、1800年の時点で、キュヴィエは、地層が深くなればなるほど、化石が現存する種と似ていないことを指摘している。
- 生物のグループ間で類似していることは、近縁性の証拠と考えられ、それは進化の変化を示唆するものであった。 ダーウィンの知的先達は、生物間の進化的関係の考えを受け入れたが、進化がどのように起こるかについて満足な説明をすることができなかった。
- この中で最も有名なのはラマルクである。 1801年、彼は生物群間の物理的類似性の説明として有機的進化を提案し、後天的特性の遺伝に基づく適応的変化の機構を提案した。 彼はキリンについてこう書いている。
「この動物は哺乳類の中で最も背が高く、アフリカの内陸部に生息していることが分かっているが、土壌はほとんど常に乾燥していて草木がないため、木を囓り、そこにたどり着くために絶えず体を酷使しなければならない。 この習性を長く続けた結果、その種族はみな、前脚が後脚より長くなり、首が非常に伸びて、キリンは後脚で立たずに頭を6メートルの高さまで持ち上げるようになった」
要するに、キリンの首が長くなったのは、高い葉っぱに届くために伸び続けた結果であると言うのである。 もちろんラルマルクの仮説したメカニズムは間違っていたが、彼の例は1800年代初頭に自然科学者が進化的変化の可能性について考えていたことを明らかにしている。
- ダーウィンは調査船ビーグル号で自然主義者として若き日に航海した際の観察から影響を受けた。 ガラパゴス諸島で彼は、異なる島のカメを識別できるようにするわずかな変化に気づきました。 また、「ダーウィンフィンチ」と呼ばれるフィンチ類も、島によって微妙な違いがあることを観察した。 さらに、これらのフィンチはすべて、600マイル東にあるエクアドル本土のフィンチと似ているようで、違うのだ。 このように、生物の分布や類似性のパターンは、ダーウィンの思考に重要な影響を与えたのである。 このページの一番上にある写真は、ダーウィン自身が『研究日誌』に掲載したフィンチのスケッチである。
- 1859年、ダーウィンは有名な『自然淘汰による種の起源』を出版した。 この本の出版は騒動となり、発売日にすべての本が売れました。 ダーウィンの思想は、宗教界や科学界からも反発を受けた。 しかし、ほとんどの科学者は、ダーウィンの主張の威力を認めていた。 今日でも、教育委員会は科学カリキュラムにおけるダーウィンの理論の有効性と適合性について議論しており、この論争をめぐってさまざまな議論が展開されている(現在進行中の対話はWWWサイトTalk.Originsを参照)。 私たちにはダーウィンの証拠や議論をすべて網羅する時間はないが、核となる考え方を検討することはできる。 この進化論は何を言っているのでしょうか?
ダーウィンの理論
ダーウィンの進化論は次のような基本的な考えを内包している。 最初の3つの考えは、ダーウィンが研究を始めたとき、すでに「種の問題」に取り組んでいた先行・同時代の自然主義者の間で議論されていたものである。 ダーウィンが最初に貢献したのは、自然淘汰のメカニズムと、進化的変化に関する多くの資料による大量の証拠である。 また、生命の歴史や現代の生物学的多様性に対する我々の理解に対して、進化がもたらす結果を思慮深く説明した。
-
種(交配する生物の集団)は時間と空間の中で変化する。 現在生きている種の代表は、最近の過去に生きていた種とは異なり、現在の異なる地域の集団は、形や行動がわずかに異なっている。 このような違いは化石記録にも及んでおり、この主張の十分な裏付けとなっている。
-
すべての生物は、他の生物と共通の祖先を共有している。 時間が経つにつれて、集団は共通の祖先を持つ異なる種に分かれることがある。 十分に過去にさかのぼれば、どのような生物のペアも共通の祖先を共有している。 例えば、ヒトは約800万年前にチンパンジーと、クジラは約6000万年前に、カンガルーは1億年以上前に共通の祖先を持つ。 共通祖先とは、共通の祖先から形質を受け継いだ生物の類似性を説明するものである。
-
ダーウィンの考えでは、進化的変化は緩やかでゆっくりしたものである。 この主張は、化石記録に残る生物の漸進的変化の長いエピソードと、ダーウィンの時代に新種の突然の出現を観察した博物学者はいなかったという事実によって裏付けされた。 それ以来、生物学者や古生物学者は、系統内の進化的変化の速度が遅いものから速いものまで、幅広く記録している。
経時変化の主要なメカニズムは、以下に詳述する自然淘汰である。 この機構は系統内の生物の特性(形質)に世代から世代への変化を引き起こす。
自然淘汰のプロセス
ダーウィンの自然淘汰のプロセスには、4つの要素がある
- 変異。 生物は(集団の中で)外見や行動に個体差を示す。 これらの変異は体の大きさ、髪の色、顔の模様、声の性質、あるいは子孫の数などに関係することがある。 一方、脊椎動物の目の数など、個体間の変異がほとんどない形質もある。
- 遺伝。 親から子へ一貫して受け継がれる形質がある。 このような形質には遺伝性があるが、環境条件に強く影響され、遺伝性の弱い形質もある。 ほとんどの個体群では、毎年、地域の資源が支えることができる数よりも多くの子供が生まれ、資源の争奪戦が繰り広げられる。
- 生存と繁殖に差異がある。 地域資源の争奪に適した形質を持つ個体は、次の世代に多くの子孫を残す。
ある世代から次の世代へ、資源をめぐる闘争(ダーウィンは「生存競争」と呼んだ)は、ある変異を持つ個体が他の個体より有利となり、それによって集団内の形質の頻度が変化する。 この過程が自然淘汰である。 より多くの子孫を残す個体に有利な形質を適応と呼ぶ。
自然選択が形質に作用するためには、その形質が遺伝的変異を持ち、資源獲得競争において有利であることが必要である。 これらの要件のいずれかが起こらなければ、その形質には自然淘汰が起こらない。 (このような形質は、ダーウィンの時代以降に発見された他の進化的メカニズムによって変化する可能性があることが分かっている)。
自然淘汰は比較優位によって作動するのであって、設計の絶対的な基準ではない。 「自然淘汰は資源をめぐる競争によって作用するため、それぞれの国の住民を、その仲間の完成度に関連してのみ適応させる」(Charles Darwin, On the Origin of Species, 1859)<61><2082>20世紀には、遺伝学がダーウィンの機構と統合されて、特定の表現型に対応する遺伝子型の生存と再生産の差異として自然選択を評価できるようになった。 自然淘汰は集団内に存在する変異にしか働かない。 このような変異は、突然変異、すなわち形質に関する遺伝暗号の一部が変化することによって生じる。 突然変異は、突然変異の潜在的な利点や欠点を予見することなく、偶然に発生する。 言い換えれば、変異は、それが必要だから生じるのではありません。
自然淘汰の証拠
自然淘汰を明確にするために例を見てみよう。
工業的メラニズムは、イギリスの70種以上の蛾に影響を与えた現象である。 最もよく研究されているのは、コショウガ(Biston betularia)である。 1800年以前は、この種の典型的な蛾は明るい模様をしていた(図2参照)。 濃い色の蛾やメラニン色の蛾は珍しく、そのためコレクターズ・アイテムとなっていた。

Figure 2.
産業革命の時代、煤煙などの産業廃棄物が木の幹を黒くし、地衣類を枯死させた。 そのため、明るい色の蛾は珍しくなり、暗い色の蛾が多くなった。 1819年に初めてメラニン色素を持つ形態が確認され、1886年にははるかに多く見られるようになり、急速な進化を遂げたことがわかる。 
やがて明るい形態は工業地帯から遠く離れた一部の地域にしか見られなくなった。 この変化の原因は、蛾のカモフラージュカラーを好む鳥による選択的な捕食と考えられています。
1950年代には、生物学者のケトルウェルが両方の形態を使って放流-再捕獲実験を行いました。 彼の結果を簡単にまとめると以下のようになります。 ブラインドから鳥の捕食を観察することで、蛾の目立ち方が食べられる確率に大きく影響することが確認されました。
捕獲成功率
|
dark moth |
||
|
non-industrial woods |
14.6 % |
4.7 % |
|
工業用材 |
13 % |
27.5 % |
局所適応-さらなる例
今日の講義ではこれまで、自然選択が進化論の基礎にあることを強調してきました。 それは適応的な変化のメカニズムを提供するものです。 環境の変化(例えば、ねぐらの木の幹の色が変わるなど)は、局所的な適応につながる可能性が高いのです。 広範囲に分布する個体群は、その範囲内の異なる場所で異なる環境条件を経験する可能性が高い。 その結果、その集団はやがて、わずかに、あるいはかなり異なるいくつもの下位集団から構成されるようになる。
以下は、集団の地域的条件への適応を説明する例である。
- ラット・スネーク(Elaphe obsoleta)は、北アメリカ東部の地域によって、明らかに異なる個体群を持っている(図3参照)。 これらは地理的に「種」と呼ぶべきか、亜種と呼ぶべきかは議論が分かれるところである。 ただし、隣接する個体群の間で交配が起こり、共通の遺伝子プールを共有することになるため、これらの個体群はすべて1つの種を構成している(種分化の講義を参照)。
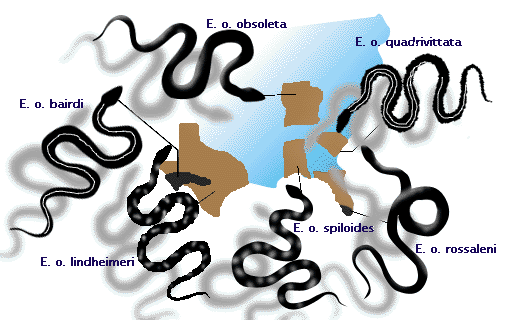
Figure 3: Elaphe obsoletaというネズミヘビの亜種で、範囲が接するところで交配しています。
- ダーウィンの航海で有名なのが、ガラパゴスフィンチです。 ダーウィンが訪れたガラパゴス諸島の各島には、世界のどこにもいない独自の種類のフィンチ(全部で14種類)が生息していました。 大きな種子を食べるのに適したくちばし、小さな種子を食べるのに適したくちばし、芽や果実を食べるオウムのようなくちばし、小さな昆虫を食べるのに適した細いくちばしのものがあった(図4参照)。 また、キツツキのように、とげを使って木の中の幼虫を探るものもいた。 (このように生態学的な役割分担(ニッチ)が多様化することが、複数の種が共存するために必要だと考えられている(この点については、後の講義で考察する)。 ダーウィンには、南米大陸から600マイルほど東に位置するフィンチの原種から、それぞれが少しずつ変化しているように見えた。 適応放散によってこれほど多くの種が形成されたのは、他の鳥類が少ないか不在で、埋めるべきニッチが空いていたことと、ガラパゴスの多数の島が地理的に隔離するのに十分な機会を提供してくれたからであろうと思われる。
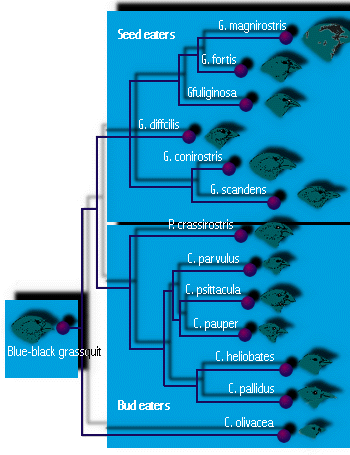
図4
安定化選択、方向性選択、多様化選択
最後に、選択について統計的に考える方法を見てみよう。 各集団がある形質–たとえばくちばしの大きさ–の度数分布として描かれるとする。 形質における変異は進化が起こるための重要な原料であることにもう一度注意してください。
次の世代では頻度分布はどのようになるのだろうか。
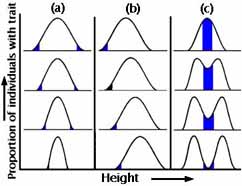
図5a~c
第1に、形質の各値(くちばしの大きさや体重)を持つ個体の割合は全く同じかもしれない。 第二に、一方向だけの変化があるかもしれない。 第三に、(その存在に議論の余地があるほど稀ではあるが)両方向の変化が同時に起こるかもしれない(例えば、くちばしが大きいものと小さいものの両方が好まれ、中間のサイズのものは犠牲にされる)。 図5a〜cは、このような自然淘汰の3つの主要なカテゴリーを捉えたものである。
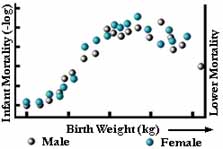
図6
安定化選択では、頻度分布の両端から極端な品種が排除される。 頻度分布は前の世代と全く同じになる(図5a参照)。 おそらくこれが最も一般的な自然選択であり、私たちはこれを無選択と勘違いしていることが多い。 実際の例としては、人間の赤ちゃんの出生時の体重がある(図6参照)。
方向淘汰では、くちばしの大きさの分布の一方の端にいる個体が特によく働くので、後続世代における形質の頻度分布は親世代におけるものからずれる(図5b参照)。 これが、私たちが通常考えている自然淘汰である。 工業的メラニズムはそのような例であった。

図7
ウマの化石系統は、方向性継承の顕著な実証を提供している。 その系統は非常に複雑で、始新世初期の小さな夜明けの馬ヒラコテリウムから、今日のおなじみのエクウスに至るという単純なものではないのだ。 しかし、全体的に見れば、ウマは森林や雑木林を移動するために作られた小さな体の祖先から、広い草原でスピードを出すために作られた長い脚を持つ子孫へと進化してきたのである。 この進化には、歯、脚の長さ、つま先の構造などの変化がよく知られている(図7参照)。
多様化(破壊的)選択のもとでは、中間的な品種を犠牲にして両極端が好まれる(図5cを参照)。 これは珍しいことだが、地理的な隔離を伴わない種形成のメカニズムを示唆しており、理論的に興味深い(種分化の講義を参照)。
まとめ
ダーウィンの進化論は、自然界に関するそれまでの考え方に疑問を投げかけ始めた思想の高まりの上に成り立っているが、将来の科学的思考の方向を根本的に変えてしまった。
ダーウィンの理論の中核は自然選択であり、連続する世代にわたって起こるプロセスで、遺伝子型の差動再生産と定義される。
自然選択には、与えられた形質における遺伝的変異と、その形質の所有に関連した生存と繁殖の差異が必要である。
自然淘汰の例は、観察と化石の記録の両方によって、十分に文書化されている。
選択は形質の頻度に作用し、安定化選択、方向性選択、または多様化選択の形をとることができる。
推薦図書
o ダーウィン,C. 1959. 自然淘汰による種の起源、あるいは生命のための闘いにおける有利な人種の保存について。 ロンドン.
o Futuyma, D.J. 1986. 進化生物学. サンダーランド、マサチューセッツ州。
o Dawkins, R. 1989. 利己的な遺伝子. オックスフォード。 オックスフォード大学出版局.
.