
















































このイラストは、印刷可能なモノクロのカラーシートとして提供されています。 ちょうどここをクリックし、新しいウィンドウで開き、コンピュータに保存する画像を右クリックします
名前。 C.megalodon(ビッグ・トゥース)
Phonetic: Meg-ah-low-don.
Named By: Louis Agassiz – 1843.
同義語: Procarcharodon megalodon、Megaselachus megalodon.
分類:Procarcharodon megalodon、Megaselachus megalodon.
Classification: 脊索動物門軟骨魚類 ラムニiformes.
種: カルチャロドンメガロドン、カルチャロクレスメガロドン、プロカルチャロドンメガロドン、あるいはオトダスメガロドンは、完全な説明のためのテキストを参照してください。 肉食。
大きさ。 多くの推定値が存在するが、通常は全長15〜17m。
知られている場所。
時代:世界各地。
化石表現:後期漸新世から前期更新世。
歯と椎骨-メガロドンの化石
歯は、C.メガロドンの最も一般的な遺跡で、大きい歯は高さが18センチメートルにもなる。 C.メガロドンの歯をいつから集めていたかは不明だが、サメの歯として科学的に認められたのは、1667年になってから。 これ以前の人々は一般的に、正直に言うとC.を思っていた。8198>
C.メガロドンの歯は、三角形で鋸歯状という表面的な類似性から、通常ホオジロザメの歯と比較されます。このような類似性、特に鋸歯から、C.megalodonはホホジロザメと同じ噛み方をしていたと考えられている。
同じ個体のC.megalodonの歯の完全セットに近いものが一緒に発見されたこともある。C.メガロドンは、他のサメと同様に、口の前方に大きな歯があり、前方に行くほど、三角形の歯が多くなり、後方に行くほど、内側に曲がっています。 この歯が獲物の肉に食い込み、前歯で切りながら逃げにくくするのです。
C.メガロドンについてよく言われる神話のひとつに、歯だけしか知られていないというものがあります。 実際には、C.メガロドンの化石は歯が圧倒的に多いのですが、脊椎動物も少しずつ増えています。 一般にサメの生態では、骨格は常に「柔らかい」軟骨でできていますが、脊椎動物は硬い石灰化軟骨からできているのです。

サイズ-メガロドンの上限サイズの推定
C.E.M.の歯が発見されて以来、メガロドンの上限サイズを推定することができる。 メガロドンの歯がサメの歯だとわかってから、ほとんどの人はすぐに「でも、サメはどのくらい大きかったのですか」と尋ねました。 正直なところ、誰かが巻尺を取れるような完全な標本がないため、正確なことはわからないのです。その代わりに、古生物学者や魚類学者は、持っているもの、この場合は歯で仕事をしなければなりません。 歯の分析に基づいてサメの長さを推定する研究は数多くありますが、最もよく言及されているのはこの4つです。
かつてメガロドンの長さを測るために使われた方法のひとつが、歯のエナメル質の高さを測ることでした。 これはジョン E. が開発したもので、この方法では、サメの体長は測定できません。しかし、この方法は、ホホジロザメの歯を比較することから始まったもので、一見すると似ているが、実はC.メガロドンの歯とはかなり異なっているため、広く科学界では正確な方法とはみなされていない。
1996年にMichael D. Gottfried、Leonard J. V. Compagno、S. Curtis Bowmanが開発した別の推定法は、歯の傾斜高さとサメの体長の間に相関関係を確立することを基本としています。この研究でも、ホオジロザメの歯の研究に基づいており、15.9mという推定値が得られた。 しかし、この方法が信頼できるとしても、与えられた歯からの推定値でしかなく、必ずしも種を特定できるわけではない
2002年、Dr.これは、歯冠の幅から顎の幅を割り出し、そこからサメの体長を割り出すという単純な原理です。 簡単に言えば、体長129cmに対して歯根が1cmあるということです。2002年には、島田健秀博士が、歯冠の比率に基づくより複雑な方法を開発した。 この方法を1996年にゴットフリードらが使用した歯に適用したところ、推定長さは15.1メートルとなった。
C. megalodonの推定体長は15-16メートルの範囲であればかなり安全と考えられ、17メートルに近い推定値は現実的に可能であると考えられている。 しかし、これらの推定値のほとんどは可能性にすぎず、現在の化石の証拠を反映していません。 それでも、15メートルという控えめな体長でも、C.メガロドンは他のどのサメも凌駕し、ホオジロザメでさえその巨大さとは比べものにならないほど、生きているか死んでいるかわかりません。
C.メガロドンがどれほどの重さだったかは、また別の研究課題ですが、その一例として、さらに幅広い推定値があります。 現在の推定値は、やはりホホジロザメとの比較に基づいており、15.9メートルのC.メガロドンの推定値は47メートルトン。 さらに大きな推定値は17メートルC.メガロドンの59メートルトン、20.3メートルC.メガロドンの103メートルトンなどとなっています。20.3mのサメは15.9mのサメの約3分の1ですが、体重は2倍以上あります。 これは、サメが長いだけでなく、筋肉量が増え、より太く頑丈になったためです。 また、サメであろうと他の動物であろうと、生息地の食料事情から、必ずそれ以上大きくなれないサイズになる。これは、体が大きければ大きいほど、燃料となる食料をより多く摂取しなければならないという論理に基づく自然な事実である。 メガロドンは、その巨大な体躯から、筋肉や肉の外層が内臓を環境から遮断し、体温を保持する「巨大熱作用」を持っていたと考えられています。 これにより代謝が高まり、より活動的になったのです。 C.megalodonmayはホオジロザメが行うことが知られているように、その頭と鼻に暖かい血液の流れをdirectingingによって巨大化のプロセスをさらに取りました。 これは、嗅覚、視覚と聴覚受容ampullaeなどのC.megalodonの脳や感覚器官は温血病代謝レベルで動作するという効果を持っているだろう。C.メガロドン歯がすべての大陸から知られているという事実は、他の成功marinepredators多くのように国際的分布を持っていたという証拠である。
C.メガロドンの全体的な外観は、その大きなサイズと固有のパワーは、それが現代のサメに比べておそらくquiteずんぐりだったことを意味しますが、最も可能性が高い今日の海で泳いでいるlamniformサメに非常に似ていただろう。
C.megalodonprobablyは、上下に動くだけでなく、体の他の部分から独立して前方に突き出すことができる可動式の上顎を持っていた。この種の顎は現代のサメにも見られ、何かを噛み砕こうとしているサメが、ただクルーズしている灰鮫とはまったく違って見えるのはこのためである。上あごを前に突き出すと、普段は口に隠れている上歯とあごが見える。 このように動くあごを持つC.メガロドンは、下あごで獲物を固定し、頭を左右に振って歯型が肉の塊を切り落とすようにするのである。歯が発見されると、C.メガロドンがどの程度の力で噛み砕くことができるかを推定することが重要になります。2008年、ホホジロザメの咬合力を推定する生体力学コンピューターモデルが作成されました。 このモデルをC.メガロドンに適用したところ、15.9メートルと大型のC.メガロドンは、108, 514ニュートン(11メートルトン強)の咬合力を発揮することが明らかにされました。また、C.メガロドンの推定サイズの上限である20.3mのC.メガロドンの咬合力を推定したところ、182, 201ニュートンと18.5トン以上でした。 このことから、C.メガロドンの推定値が低くても、C.メガロドンの咬合力は低いことがわかりました。 もうひとつ考慮すべきは、C.メガロドンはおそらく、同じような歯列を持つ他のサメと同様に、頭を左右に振っていたという事実だ。 つまり、C.メガロドンの不幸な獲物にかかる実際の力は、おそらくさらに大きいということである
一方、C. メガロドンの研究の大半は、そのようなことはない。 しかし、C.メガロドンの研究の中心は歯であり、見落としがちなのが脊椎骨です。 脊椎骨は数が少ないため、歯のように誰もが研究できるわけではありませんが、脊椎骨には明らかに同心円が見られます。 この同心円は、木の切り株に見られる成長輪と本質的に同じで、現在海を泳ぐ他の現代のサメでも見ることができます。この同心円を数えることで、化石から知られるC.メガロドンの年齢を25~40歳と推定しており、古生物学者はC.メガロドンがこれよりもさらに長く生きられた可能性を示唆している。 メガロドンは生きたまま子供を産んだと考えられている。 しかし、C. megalodondがどのようにこれを行ったかについては、サメには2つの方法があるため、まだ議論の余地がある。 1つ目は、胎生と呼ばれ、子ザメが母親の中で成長し、生まれる準備をする方法。 この方法で生まれる子ザメは、へその緒を通して栄養を与えられ、子ザメが生まれると通常はそのまま胎盤が放出される。
2つ目の方法は卵胎生で、卵の中で成長する。 ただし、卵胎生を示すサメは卵を産まず、母サメの体内に保持する。 母サメとへその緒で結ばれず、卵黄を消費して成長する。C.メガロドンの仔は通常、尾から先に出てくるように描かれており、これが通常の方法と考えられているが、現在のサメの中には頭から先に生まれるものもいる。
現在のサメと同様に、C.メガロドンの仔も頭から先に生まれている。 メガロドンは、おそらくどこでも産めるわけではなく、保育所と呼ばれる場所を選んで産んだ。 保育所とは、幼いサメが他の大型捕食者から守られながら生活し、狩りができる場所。 保育所にいるサメは、魚、頭足類(タコやイカなど)、カメなど、あらゆる生き物を攻撃して食べる総合生物と考えられている。
潜在的な保育園を識別するために古生物学者は、小さなC.のforconcentrationsを見る。 これらの歯でbeplentifulと思われるOneareaは中央アメリカと米国の最も南のareaofです。 当時の高い海面がofthe領域の多くを水没させたため、漸新世に向かって戻って、パナマは存在しませんでした。 この地域は中米海峡として知られており、太平洋と大西洋の間に海のパスを形成していた。

Fossil evidence strongly suggeststhat during the Miocene between North and SouthAmerica had a very high population of C. megalodonpups. また、クジラ(紫)などの初期の鯨類は、当時中米海峡と呼ばれていた海域を太平洋と大西洋の間の通路として利用していたことが化石からわかっています。 火山活動の活発化と海水準の低下により、この海路は閉鎖され、餌の減少や海流が劇的に変化した。 8198>
歯の大きさを調べてみると、保育所にいたメガロドンは体長2~3メートルと小さいことがわかりました。 しかし、これは生まれたときから大きかったわけではなく、その地域で活動していた幼生の歯であることを意味しています。このように、C.メガロドンは成長するにつれて大きくなり、やがて浅瀬から外洋に出ることになります。 このとき、大型の海洋生物を専門に攻撃しなければならない第二段階が始まります。
Prey items – What did megalodon eat?
「C.megałodond」は恐竜を食べたとする説。 C. megalodonが恐竜を食べることは不可能である(本当に恐竜や大型海洋爬虫類を食べることができたサメは、Cretoxyrhinaを調べてほしい)。 C. megalodonの好物は、鯨類、特に小型から中型の鯨だったようだ。 また、C. megalodonの好物は、鯨のような小型の鯨だったという証拠も残っている。しかし、C.megalodonが何を狩ったかは、個体の年齢によって異なり、小さいC.megalodonはugongなどの動物を狩り、大きいC.megalodonは大きいクジラを狩りました。 鯨類の脊椎骨には、下方からの突然の衝撃によって引き起こされたと解釈される圧縮損傷が見られる。 これにより、C.メガロドンがターゲットに見つからないように下方から鯨に接近するシナリオを再構築することができた。もし鯨がC.megalodonの顎の中に入らなければ、鯨は衝撃で気絶し、C.megalodonに噛み付く時間ができたと考えられます。しかし、少なくとも1つの脊椎骨の化石は、このような攻撃を受けたにもかかわらず、治癒したことが知られています。 これは、この場合、幸運な動物は単に攻撃を生き延びただけでなく、傷が治癒するのに十分長く生きたことを示しています。

Special note* – 上記のシーケンスは、C. megalodon鮫が獲物を狩った一つの可能性を説明することを意図したものです。 その結果、C.メガロドンは肋骨のような骨のある部分を狙っていたことがわかった。 ここでは、簡単に折れない非常に頑丈な歯と、簡単に骨を折ることができ、その結果、守るべき内臓に大規模な損傷を与える破砕咬合力の2つが有利に働いていたのだ。この方法に対するさらなる支持は、歯が鈍化した部分の圧縮骨折から得られ、骨のような硬い物質との強い衝撃を示唆している。
肋骨などの部位に噛み付くには大きすぎる可能性がある大型鯨を攻撃するとき、C. megalodonは戦術を変更した。 重要臓器を狙う代わりに、獲物を固定化しようと尾部を攻撃したのである。サメは呼吸をするために常に前方に泳いでいるが、猛烈なスピードは短時間しか維持できない。 これは、サメの白筋(全筋肉の約90%)は急激なスピードを出すために使われ、すぐに疲れるが、赤筋(同約10%)は力は弱いが信じられないほどの耐久性があるため、赤筋が通常の巡行に使われるためである。このように、C.メガロドンは、大きな鯨を不自由にすることで、無理をせず、ゆっくりと餌を食べることができるのです。
C.メガロドンは、単に狩りをするには大きすぎ、スカベンジャーだったのではないか、という主張もある。 多くの大型鯨類の歯型だけでなく、傷を示す圧倒的な化石証拠を前に、この主張はありえないどころか、ほとんど不可能だと考えられているのである。サメや肉食動物の多くは死体から餌をとるが、それが排他的なスカベンジャーになるわけではない。 また、ただ食べるだけで生きている海洋動物は、死んだ動物が海の底に沈むのを待つボトムフィーダーになる傾向がある。 C.メガロドンのサイズを推定すると、それを維持するためにどれくらいの餌が必要かも推定できるようになった。これは、スカベンジャーにとっては途方もない量である。 化石の証拠、生体モデル、サメのライフスタイルや生態に関する知識をすべて考慮すると、スカベンジャーがC.megalodonの生存方法である可能性は最も低いという結果になる。
絶滅-なぜメガロドンは消えたのか?
180万年前の更新世第1期(ゲラシアン)の終わり頃にC.megalodondisは化石記録から消えている。 この消滅は、C.メガロドンが着実に減少していることが特徴的である。 C.メガロドンが絶滅した理由についてはいくつかの説があるが、1つの事象だけでなく、一連の変化により滅亡した可能性が高いと思われる。
C.メガロドン絶滅の引き金となったのは、地球規模の冷却だったようです。 そもそも、C.メガロドンが巨大な熱量によって温血病的な代謝を行っていたとすれば、同サイズの完全に冷血な生物よりも高い摂取カロリーを必要とします。 水の温度が低いほど、その差はより大きくなり、C.メガロドンは温度低下に対応するためにさらに多くの食物を必要としたと思われます。 また、巨大な熱は、真の温血動物の代謝の代わりにはならず、冷たい水の中のサメは、暖かい水の中にいたときよりも鈍重である可能性があり、狩りの能力をさらに妨げるものであった。そのため、このような「曖昧さ」は、「曖昧さ」そのものであり、「曖昧さ」である。パナマ地峡の形成の結果、中米海路が閉鎖され、鯨の化石が大量に発見されたことから、鯨の重要な移動ルートとして使用されていたようである。中新世には20種以上のクジラが生息していたが、現在では6種しかいない。 残りのクジラはまだ移動していたが、ヒゲクジラが食べるのに適した無脊椎動物がより豊富なため、おそらく極域を好んだと思われる。その数はマッコウクジラでalsoodramatically減少したように歯鯨はどちらか実行可能なオプションであったようです今日まで生き残った唯一の大型tothewhale。 C. megalodonrestricted to the warmerocean waters it longer had constant year round access to the foodsupply that was most adapted to kill.このように、ヒゲクジラは、それが殺されるように適応されていた。
C.メガロドンの巨大なサイズは、利用可能な唯一の他の獲物が小さく、高速で、たとえ捕獲されたとしても、大きなクジラがやったように栄養の同じレベルを提供しなかったとして、これらの時代にworkagainstit疑いもなくです。また、メガロドンの生存戦略として共食いが提案されているが、これは食べるべき他のメガロドンがいる限り有効である。 もし実際にそうなれば、共食いはメガロドンの数をさらに減らし、その結果、繁殖する数が制限される。
これに関連して、海面の変化による保育場所の喪失の可能性もある。パナマ地峡の形成は、C.メガロドンの稚魚の歯が大量に出土したことから、そのような育児場所を取り除いたと思われます。 育児場所が失われたことは、C.メガロドンの稚魚自身が捕食されやすくなり、おそらく、生き残りをかけて他のC.メガロドンをも捕食したであろうことを意味しています。
最後の説は、新しい捕食者の台頭で、特に今日シャチとして知られているオルカに代表される猛禽類の進化に言及する。 C.メガロドンの数が減少するにつれ、デルフィニドの数が増加したのである。しかし、これらの新しい捕食者の台頭がC.メガロドンの衰退の一部を再生したかどうかを言うことは困難であるとして、それは同様に新しいpredatorsroomがthrive.There間の捕食者と餌生物の相互作用を示す化石の証拠を許容するC.メガロドンが減少していた可能性があります。
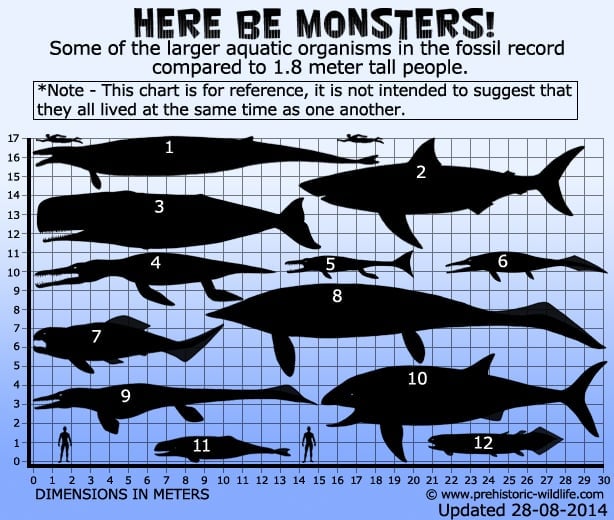
1 – バシロサウルス(クジラ)、2 – C.megalodon – 平均下推定(サメ)、3 – Livyatanmelvillei – 下推定(クジラ)、4 – Pliosaurusfunkei, a.k.s.
2 – C.megalodon – 下推定(サメ)、3 – Pliosaurusfunkei – 上推定(クジラ)、4 – C.megalodon – 下推定(サメ)。プレデターX(プリオサウルス)、5-プレシオサウルス(タラトサウルス)、6-タラトアルコン(魚竜)、7-ダンクレオステウス(節足動物のプラコダーム)、8-シャスタサウルス(魚竜)、9-ティロサウルス(蚊竜)、10-リーディヒシス-上位推定(魚)、11-ブリグモフィゼーター(鯨)、12-リゾフス(lobe finned fish).
最後の生き残り?
C.メガロドンは更新世を生き残り、完新世まで海を泳いでいたという説があります。 その根拠は、1872年にHMSチャレンジャーの乗員によって発見され、1959年に検査したところ1万年しか経っていないと考えられた歯の部分から来るそうです。しかし、このテストは化石に付着した二酸化マンガンのレベルを測定するもので、同じ時代のものでも化石によって付着する二酸化マンガンのレベルが異なるため、今では欠陥のある方法と考えられています。 この歯を後の電波炭素年代測定法にかけたところ、窒素レベルが低すぎてテストできないことが判明し、この歯はテスト不可能とされ、C. megalodonが更新世初期に絶滅したという以前の推定結果は有効なままになっています。

分類-メガロドンはホオジロザメと関係があるか?
おそらく、C. megalodonに関する最大の論争点は、それが実際に今日我々が知っているホオジロザメと関係があるかどうかである。 C. megalodonの比較は以下の通りです。 メガロドンとホホジロザメの比較は、ほとんどが、メガロドンが最大のサメであり、ホホジロザメが現在知られている最大のサメであるという原則からきている。 また、両者の歯はしばしばほぼ類似していると見られている。C.メガロドンをカルチャロドンに含める支持者は、歯の類似性を、C.メガロドンとホホジロザメの両方がパラオカルカロドンの子孫であることの結果であると指摘しています。 また、C.メガロドンとホオジロザメの歯は、C.メガロドンの歯よりずっと細く、より優美である。 また、C.メガロドンが、ホオジロザメの歯よりずっと細い。また、C.メガロドンの歯はホオジロザメと全体的に似ていると考えられているが、C.メガロドンの第3前歯(上顎の中心から3番目の前歯)は、最初の2本と同様に下を向いており、ホオジロザメとは異なる形をしている。 C.megalodon 前歯にはまた、ホオジロザメにはないシェブロン状の傷があり、これは歯の冠と根の間の特徴的なものだ。
C.メガロドンをカルチャロドン属に分類する代わりに、より古いカルチャロクレス属に分類することも考えられる。 この分類の主な論拠は、Carcharoclesauriculatusという別の古代大型鮫が、実はC.メガロドンの祖先と考えられていることである。 C.メガロドンの18cmの歯に換算すると、C.メガロドンより3分の1ほど小さくなる。 それでも、Carcharoclesauriculatusが大きくなり、C. megalodonが生まれた可能性は十分にあります。
このようにC. megalodonがCarcharoclesに含まれると、実際には、サメが歯の側枝を失う移行を完了させることになります。この変遷は、Otodusobliquusに明確に存在する側歯尖に始まり、Carcharoclesauriculatusでは歯尖が減少し、C. megalodonでは歯尖がない。これらの歯にはまた、歯冠と歯根の間にシェブロン形の傷があるが、これはオオシロ歯にはないことである。
また、C.メガロドンはホオジロザメの祖先であり、時間の経過とともにホオジロザメは単に小さくなったという説もある。 しかし、この考え方の最大の問題は、ホオジロザメがC.メガロドンよりずっと前に実際に海で泳いでいたことである。 メガロドンは絶滅し、ホホジロザメの歯の化石はメガロドンが絶滅する1400万年以上前の1600万年前の中新世にまでさかのぼる。 この説の支持者は、ホホジロザメがメガロドンの小型種から進化した可能性をまだ主張している。 サーベルネコのスミロドンをご存知の方は、同じ祖先から生まれたと思われる全く異なる3つの種が、同じ時代に一緒に活動していたことをご存じかもしれません。しかし、C.メガロドンからホオジロザメへの変化については、歯が小さくなるだけではなく、ホオジロザメの形に着実に変化していることを示す移行リンクはないようである。
2012年、C.megalodonとCarcharodoncarchariasは無関係であるという説は、Carcharodonの新種、Carcharodonhubbelli(ハベルホワイトシャーク)の記載によって、もう少し裏付けが取れました。 カルチャロドン・ハッベリの歯は、カルチャロドン・カルカリアスとアオザメを含むイスルス属をつなぐ移行型であると解釈する人もいるようだ。 メガロドンのような巨大ザメとアオザメの間に直接的なつながりはないとする研究者が多いことから、カルチャロドン・カルカリアスはメガロドンとは別物であり、ひいてはメガロドンはカルチャロクレス属に含まれるべきものであることを示唆しているのかもしれない
メガロドンとホオジロの全体の形態の類似性は、遺伝的交配の結果というよりも進化の結果として生まれた可能性が最も高い。この基本的な体型は「魚雷型」と呼ばれ、前方が尖って中央が広くなり、後方が細くなっている。 この形は自然界で何度も繰り返されており、単に水中移動に最も適した形であるため、ジャストシャークに特有の形ではないことは確かである。 この考え方は、オトドス型の歯から後世のメガロドン型の歯への移行が目に見えるという研究結果によるものである。
Further reading
– Recherches sur les poissons fossiles/par Louis Agassiz – Neuchatel:Petitpierre.p. 41.Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: Theory: They: Theory: Theory: They: Theory: They: Theory: They: They: They: They: No. – Louis Agassiz – 1833-1843.
– Size of the Great White Shark (Carcharodon) – Science Magazine 181(4095): 169-170 – John Randall – 1973.
– Carcharodon megalodon from the Upper Miocene ofDenmark, withcomments on elasmobranch tooth enameloid: coronoi’n – Bulletin of theGeological Society of Denmark (Copenhagen: Geologisk Museum) 32: 1-32.
-2000.
-3000.- Svend Erik Bendix-Almgreen – 1983.
– Catalogue of Cuban fossil Elasmobranchii (Paleocene to Pliocene) andpaleogeographic implications of their Lower to Middle Mioceneoccurrence – Boletín de la Sociedad Jamaicana de Geología (Cuba) 31:7-21 -M. Iturralde-Vinent, G. Hubbel & R. Rojas – 1996.
– The Megatooth shark, Carcharodon megalodon: Roughtooted, hugetooted – Mundo Marino Revista Internacional de Vida (non-refereed)(Marina) 5: 6-11. – J. C. Bruner – 1997.
– ジャマイカ産サメ化石 – 瑞浪市化石博物館紀要.pp. 211-215. – Stephen Donovan & Gunter Gavin – 2001.
– ニュージーランド後期漸新世のCarcharodon angustidens (Chondrichthyes,Lamnidae) の関連標本、Carcharodon interrelationshipsに関するコメント – Journal ofVertebrate Paleontology 21(4): 730-739. – M. D. Gottfried & R. E. Fordyce – 2001.
– ヨシキリザメの歯サイズと体長の関係 – 化石研究(日本)35 (2): 28-33. – 島田健秀 – 2002.
– プエルトリコの中期中新世に生息するラムニッドシャークCarcharodon megalodonfの新記録 – Caribbean Journal of Science 39: 223-227. Böhme – 2003.
– Carcharocles megalodon (Lamniformes:Otodontidae) の年齢:層序学的記録のレビュー – 日本古生物学会 (PSJ) 75 (75): 7-15. – 河辺秀夫・後藤正俊 &金子直知 – 2004.
– ベネズエラ新第三紀のオオテンジクザメと広歯類(Lamnidae): カリブ海の浅海性魚類群集における役割 – Caribbean Journal of Science 40 (3): 362-368. – O.Aguilera & E. R. D. Aguilera – 2004.
– ホオジロザメの祖先を探る -Journal of VertebratePaleontology 26 (4): 806-814 – K. G. Nyberg, C. N. Ciampaglio &G. A. Wray – 2006.
– Late Neogene Oceanographic Change along Florida’s West Coast:Evidence and Mechanisms – The Journal of Geology (USA: The Universityof Chicago) 104 (2): 143-162. – また、このような環境下において、「環境問題」を解決するためには、「環境問題」を解決するために、「環境問題」を解決するために、「環境問題」を解決する必要があります。 – ジャーナル・オブ・ズーロジー 276 (4): 336-342. -S. 塚田晋一郎, 塚田晋一郎, 塚田晋一郎, 塚田晋一郎, 塚田晋一郎 – 2008.
– カリアコウのケンディエース層とグランドベイ層における中新世のサメ – カリブ海科学誌(2008). 44 (3)pp. 279-286. – Roger Portell, Gordon Hubell, Stephen Donovan, JeremyGreen, David Harper & Ron Pickerill – 2008.
– Giant-toothed white shark and cetacean trophic interaction from thePliocene Caribbean Paraguaná Formation – Paläontologische Zeitschrift (Springer Berlin) 82 (2): 204-208. – また、このような研究成果を踏まえて、本学科の研究者・大学院生を対象とした研究報告会を開催しました。 McFadden & GordonHubbell – 2010.
– The Great White Shark Carcharodon carcharias(Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic -Revista Española de Paleontología (Portugal) 25 (1): 1-6. – MiguelTelle Antunes, Ausenda Cáceres Balbino – 2010.
– Patterns and ecosystem consequences of shark declines in the ocean -Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055-1071. -Francesco Ferretti, Boris Worm, Gregory L. Britten, Michael J. Heithaus& Heike K. Lotze – 2010.
– White shark Carcharodon(Lamniformes: Lamnidae)Origin based on recalibration of upper Neogene Pisco Formation of Peru- Palaeontology 55(6):1139-1153 – D. J…また、このような環境下での研究開発には、研究者だけでなく、研究者間の情報交換の場が必要である。 – また、このような研究成果を踏まえて、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学術振興会特別研究員奨励賞」、「日本学士院会員奨励賞」、「日本学士院会員奨励賞」を受賞しました。 Ehret,Austin J. W. Hendy, Bruce J. MacFadden & Carlos Jaramillo -2013.
– Carcharocles megalodonはいつ絶滅したのか? ANew Analysis of theFossil Record. – PLOS ONE. 9 (10): e111086. – C. Pimiento & C.F. Clements – 2014.
– 絶滅した巨大鮫Carcharoclesmegalodonの体格傾向:海洋頂部捕食者の深層時間的展望. – 古生物学. 41 (3):479-490. – C. Pimiento & M. A. Balk – 2015.
– EasternGuadalquivir Basin(Upper Miocene, South Spain)におけるCarcharocles megalodonの記録. Estudios Geológicos. 71 (2): e032. – M.Reolid & J. M. Molina – 2015.
– Carcharocles megalodonovertimeの地理的分布パターンから絶滅メカニズムに関する手がかりを得た。 – Journal ofBiogeography. 43 (8): 1645-1655. – C. Pimiento, B. J. MacFadden, C. F.Clements, S. Varela, C. Jaramillo, J. Velez-Juarbe & B.R.Silliman – 2016.
– メガトースザメ、Otodus megalodon(Lamniformes:Otodontidae), the size of the megatooth sharkの再検討. – 歴史生物学:1-8. – 島田健秀 -2019.
– メガトゥース・シャークOtodusmegalodonの鮮新世初期の絶滅:北太平洋東部からの視点. – PeerJ. 7:e6088. -R. W. Boessenecker, D. J. Ehret, D. J. Long, M. Churchill, E. Martin& S. J. Boessenecker – 2019.
– Carcharocles chubutensis と Carcharoclesmegalodon (Otodontidae, Chondrichthyes) の移行:側頭葉が失われた時期. – を、”Journal of Vertebrate Paleontology. 38 (6): e1546732. – V. J.Perez, S.J. Godfrey, B.W. Kent, R.E. Weems & J.R. Nance -2019.
– 絶滅巨大鮫Otodus megalodonの体寸法:2Dreconstruction.J.Perez. – サイエンティフィック・リポーツ. 10 (14596): 14596. – J. A.Cooper, C. Pimiento, H. G. Ferrón & M. J. Benton -2020年.
—————————————————————
Random favorites
 |
 |
 |
 |
プライバシーポリシー&クッキーポリシー
。