真核生物の細胞分裂には、大きく分けて有糸分裂と減数分裂の2種類があります。 有糸分裂は成長や治癒のために新しい体細胞を作り出すのに使われ、減数分裂は性細胞(卵と精子)を作り出すのに使われる。 減数分裂については、後の章で説明する。
細胞周期は、細胞の成長と細胞分裂を伴う一連の秩序だった出来事であり、有糸分裂によって2つの新しい娘細胞を生み出すものである。 細胞周期の長さは、個々の生物の細胞内でも非常に多様である。 ヒトの場合、細胞のターンオーバーの頻度は、胚発生初期の数時間から、上皮細胞では平均2~5日、大脳皮質神経細胞や心筋細胞などの特殊な細胞では一生涯、細胞分裂をせずに過ごすこともある。 また、細胞が細胞周期の各段階で過ごす時間にもばらつきがある。 分裂の早い哺乳類細胞を培養(体外で最適な増殖条件下で)した場合、その周期の長さは約24時間である。 細胞周期におけるイベントのタイミングは、細胞の内部と外部の両方のメカニズムによって制御されている。
細胞分裂への道を歩む細胞は、成長、DNA複製、分裂の一連の正確なタイミングと慎重に制御された段階を経て、遺伝的に同一の二つの細胞を生成する。 細胞周期には、間期と分裂期という2つの主要な段階がある(図1)。 間期では、細胞は成長し、DNAが複製される。 分裂期には、複製されたDNAと細胞質の内容物が分離し、細胞が分裂する。
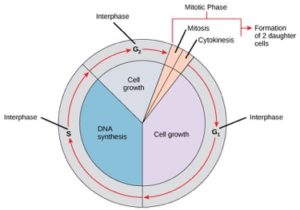
間期では、細胞は通常のプロセスを経ながら、細胞分裂の準備も行う。 細胞が間期から分裂期へ移行するためには、多くの内的および外的条件が満たされる必要がある。 間期の3つの段階をG1、S、G2と呼ぶ。
G1期(第1ギャップ)
間期の第1段階は、顕微鏡で見るとほとんど変化が見えないため、G1期(第1ギャップ)と呼ばれる。 しかし、G1期の間、細胞は生化学的なレベルではかなり活発である。
S期(DNAの合成)
間期を通じて、核DNAは半凝縮クロマチンの状態にあります。 S期には、DNAの複製が進み、同じDNA分子のペア-姉妹染色分体-が形成され、染色体の中心部にしっかりとくっつきます(図2)。 染色体はセントロメアと呼ばれる部位でくっついたままです。 Photo credit: Lisa Bartee
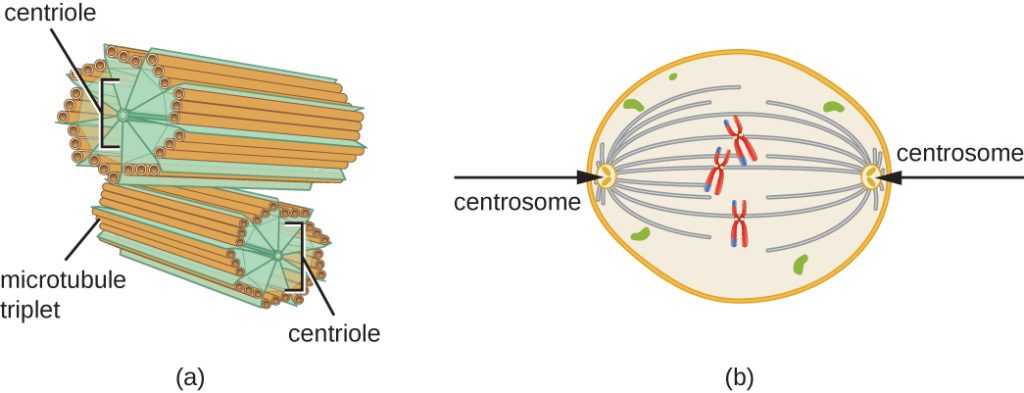
S期にはセントロゾームも複製される。 この2つの中心体は、有糸分裂の際に染色体の動きを調整する装置である有糸分裂紡錘体を生み出します。 各動物細胞の中心部には、互いに直角をなす一対の棒状の物体、中心核が付随している。 中心体は、細胞分裂を組織化するのに役立っている。 植物やほとんどの菌類など、他の真核生物の中心体には中心核は存在しない。
G2期(セカンドギャップ)
G2期では、細胞はエネルギー貯蔵量を補充し、染色体操作に必要なタンパク質を合成する。 いくつかの細胞小器官は複製され、細胞骨格は有糸分裂期のための資源を提供するために解体される。 G2期にはさらに細胞増殖が行われることもある。
有糸分裂期
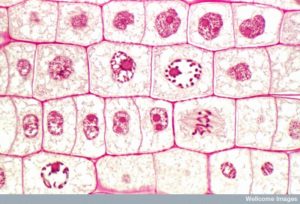
2つの娘細胞を作るには、核の中身と細胞質を分けなければなりません。 分裂期は、重複した染色体を並べ、分離し、細胞の反対側の極に移動させ、新しい同じ2つの娘細胞に分割する多段階のプロセスである。 有糸分裂期の最初の部分である有糸分裂は、5つの段階からなり、核分裂を成し遂げる(図5)。 有糸分裂期の第2部分は、細胞質成分を2つの娘細胞に物理的に分離する「サイトキネシス」と呼ばれるものである。 有糸分裂の段階はほとんどの真核生物で似ているが、植物細胞のように細胞壁を持つ真核生物では、細胞質分裂の過程はかなり異なる。
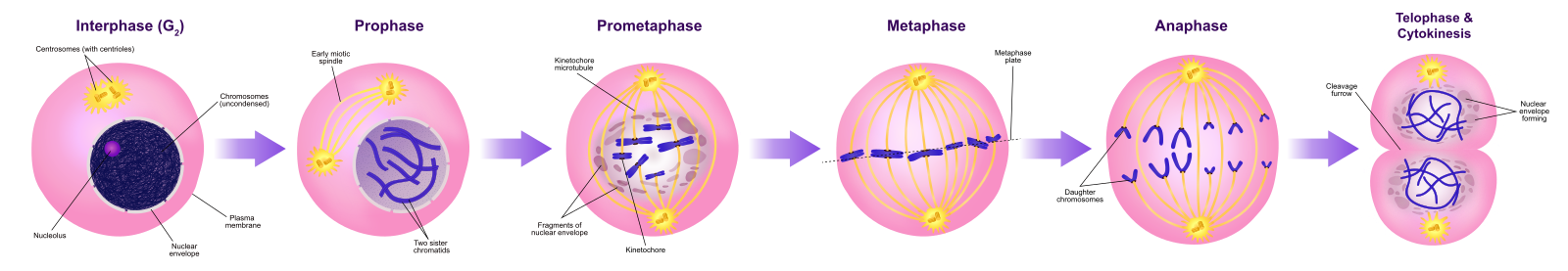
Prophase
「第一期」である前段階では、核膜が小さな小胞に分解し始め、膜小器官(ゴルジ装置や小胞体など)が断片化して細胞の端に分散していきます。 核小体は消失する。 中心体は細胞の反対側の極に移動し始める。 微小管繊維が長くなり、中心体の間を押し広げながら、分裂紡錘体を形成する。 姉妹染色分体はコンデンシンタンパク質の助けを借りて、より堅く巻き付き始め、光学顕微鏡で見えるようになる。 Photo credit Kelvin13; Wikimedia.
Prometaphase
「最初の変化期」であるプロメタフェーズでは、プロフェーズで始まった多くのプロセスが引き続き進行する。 核膜の残骸が断片化する。 微小管はさらに集合し、かつての核領域の長さを横切って伸びるので、有糸分裂紡錘体は発達し続ける。 染色体はより凝縮され、ばらばらになる。
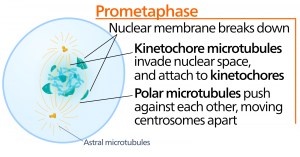
キネトコアのタンパク質は、有糸分裂紡錘体の微小管を引き寄せて結合させる。 紡錘体微小管が中心体から伸びるとき、これらの微小管の一部はキネトコアと接触し、しっかりと結合する。 分裂繊維が染色体に付着すると、染色体は姉妹染色体の動原体が反対極を向くまで方向転換される。 最終的には、すべての姉妹染色分体は、キネトコアを介して対極の微小管に結合する。 染色体と結合していない紡錘体微小管は極性微小管と呼ばれる。 これらの微小管は2極の中間で互いに重なり合い、細胞の伸長に寄与している。 アストラル微小管は極付近にあり、紡錘体の配向を助け、有糸分裂の制御に必要である。

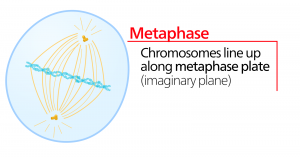
Metaphase
変化期であるメタフェースでは、すべての染色体は細胞の両極の中間にあるメタフェース板(赤道面)という平面に整列しています。 姉妹染色分体はまだコヒーシンタンパク質によって互いにしっかりと結合しています。 このとき、染色体は最大限に凝縮されています。
Anaphase
「上昇期」と呼ばれるアナフェースでは、コヒーシンタンパク質が分解し、姉妹染色分体はセントロメアで分離されます。 染色体は、微小管に結合している中心体に向かって急速に引き寄せられる。 メタフェース板で極性微小管が互いに重なり合うと、細胞は目に見えて細長くなる(楕円形)。
Telophase
「距離相」であるtelophaseでは、染色体は反対極に達し、脱凝結(ほぐれる)を始め、クロマチン配置に緩和される。 紡錘体は解重合してチューブリン単量体となり、各娘細胞の細胞骨格を構成するのに使われる。 核膜が染色体の周りに形成され、核小体が核領域内に現れる。
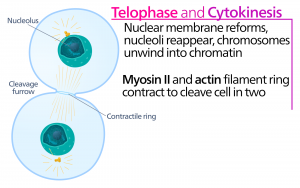
Cytokinesis
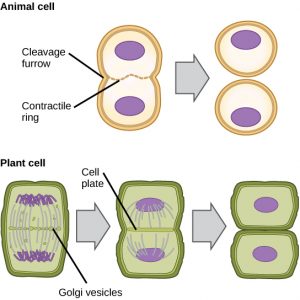
「細胞の動き」は、細胞質成分が2つの娘細胞に物理的に分離することによって細胞分裂が完了する、分裂期の第2主要段階である。 細胞の構成要素が分割され、2つの娘細胞に完全に分離されるまで、分裂は完了しない。 ほとんどの真核生物では有糸分裂の段階は似ているが、植物細胞のように細胞壁を持つ真核生物では細胞質分裂の過程はかなり異なっている。
動物細胞のように細胞壁を持たない細胞では、細胞質分裂はアナフェースの開始に続いて行われる。 アクチンフィラメントからなる収縮環は、前メタフェース板の細胞膜のすぐ内側に形成される(図12)。 アクチンフィラメントが細胞の赤道部を内側に引っ張り、亀裂を形成する。 この裂け目は「裂け目」と呼ばれる。 植物細胞では、娘細胞の間に新しい細胞壁が形成されなければならない。 間期には、ゴルジ体が酵素、構造タンパク質、グルコース分子を蓄積し、小胞に分かれて分裂する細胞全体に分散する(図12)。 テロフェーズの間、これらのゴルジ体小胞は微小管上を移動し、メタフェース板でフラグモプラスト(小胞構造体)を形成する。 そこで小胞は融合し、中心から細胞壁に向かって合体する。この構造は細胞板と呼ばれる。 さらに小胞が融合すると、細胞板は大きくなり、細胞周縁部で細胞壁と合体する。 酵素は、膜層の間に蓄積されたグルコースを使って、新しい細胞壁を作る。 ゴルジ膜は新しい細胞壁の両側の細胞膜の一部になる。
有糸分裂と細胞質分裂の概要
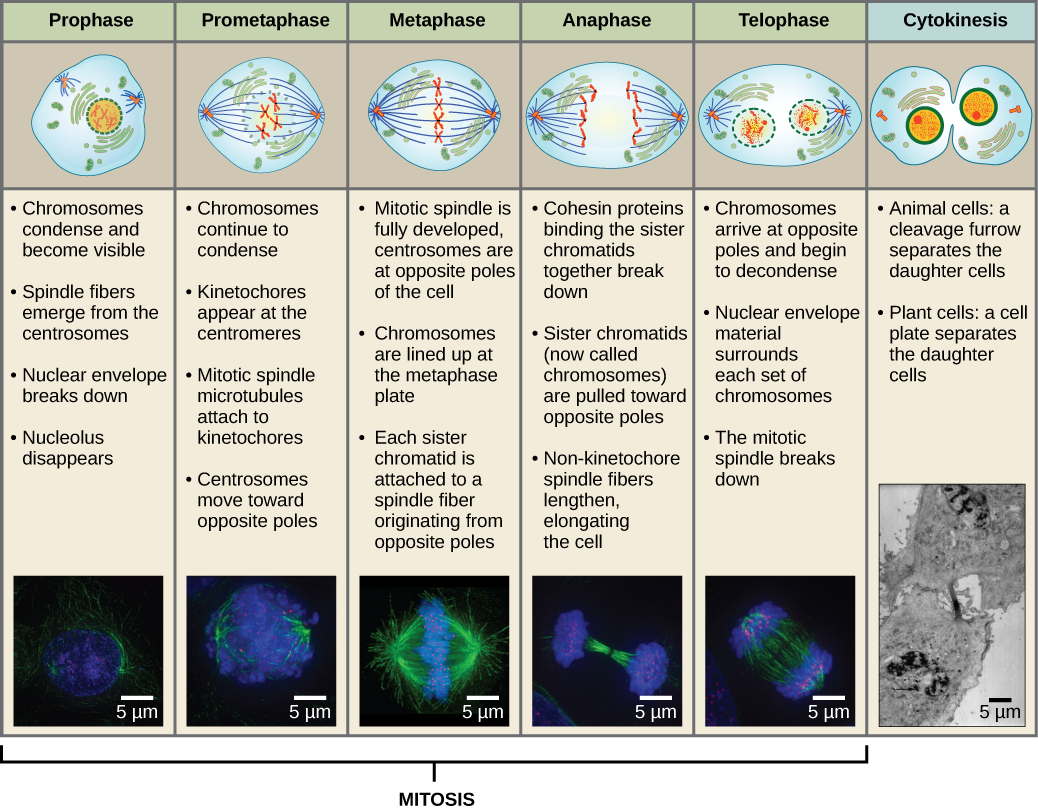
図13 有糸分裂は前段階、前周期、中期、分裂期、分裂期の5段階に分けられ、前周期と分裂期には、細胞質分裂が行われる。 下の写真は、蛍光色素で人工的に染色した細胞を蛍光顕微鏡で撮影したもので、青い蛍光はDNA(染色体)、緑の蛍光は微小管(紡錘装置)を示す。 (クレジット “mitosis drawings”: Mariana Ruiz Villarealの作品を改変、クレジット “micrographs”: Roy van Heesbeenの作品を改変、クレジット “cytokinesis micrograph”: Wadsworth Center/New York State Department of Health; scale-bar data from Matt Russell)
G0 Phase
すべての細胞が、新しくできた娘細胞がすぐに間期に入って、その後に有糸分裂期が続くという古典的な細胞周期のパターンを守っているわけではないのです。 G0期にある細胞は、活発に分裂の準備をしているわけではない。 細胞は細胞周期から外れて静止(不活性)期にある。 外部からの信号でG1が始まるまで一時的にG0に入る細胞もある。 成熟した心筋や神経細胞など、決して分裂しない、あるいはほとんど分裂しない他の細胞は、永久にG0にとどまる)
。