Introducere
Charophytes or basal Streptophytes; (Becker and Marin, 2009; Leliaert et al., 2012) constituie un sortiment taxonomic divers de alge verzi existente de apă dulce și terestre care prezintă o gamă largă de forme unicelulare, filamentoase și „parenchimatoase” (Graham, 1993; Lewis și McCourt, 2004). O descendență ancestrală de carofite a apărut pe uscat și a colonizat pământul în urmă cu 450-500 de milioane de ani. Aceste organisme s-au adaptat la condițiile terestre, au devenit capabile să supraviețuiască și să se reproducă atunci când sunt expuse complet la atmosferă, iar unii membri au evoluat în cele din urmă în plante terestre (Zhong et al., 2013; Delwiche și Cooper, 2015). Această „terestruizare” a plantelor verzi a reprezentat un eveniment biologic esențial care a schimbat pentru totdeauna biogeochimia și istoria naturală a planetei. Proliferarea ulterioară a plantelor terestre a schimbat condițiile atmosferice și a alterat și mai mult condițiile solului și a permis apariția altor forme de viață diverse pe uscat. Evoluția plantelor terestre a transformat, de asemenea, istoria omenirii, cel mai semnificativ prin inovarea agriculturii și crearea, în consecință, a civilizației umane moderne. În parte datorită semnificației lor evolutive, carofitele au primit o atenție semnificativă din partea biologilor vegetali în ultimele decenii (Pickett-Heaps și Marchant, 1972; Pickett-Heaps, 1975; Mattox și Stewart, 1984; Becker și Marin, 2009; Harholt et al., 2016). Cu toate acestea, pe măsură ce s-au adunat noi date suplimentare cu privire la biologia acestor alge, ele au devenit modele importante pentru înțelegerea fenomenelor de bază în biochimie, biologie celulară, biologie a dezvoltării, ecologie și, din ce în ce mai mult, biologie moleculară (Delwiche și Cooper, 2015; Lemieux et al., 2016). Seria „Frontiers in Plant Science” privind „Charophytes” ilustrează importanța acestor organisme în mai multe domenii specifice de cercetare în domeniul biologiei plantelor. Această mini-revizuire evidențiază atributele charophytelor ca organisme model în diverse domenii de cercetare. De asemenea, se speră că va încuraja o nouă generație de oameni de știință să extindă utilizarea acestor alge în cercetarea fundamentală și să inițieze screening-ul altor taxoni care, la rândul lor, pot duce la identificarea și utilizarea de noi charophytes model.
The „Line-Up” of Charophytes
În timpul anilor 1970 și 1980, au fost descoperite metode ultrastructurale (de ex, mecanismul de diviziune celulară, substructura aparatului flagelar) și datele biochimice (de exemplu, profilurile enzimatice) au fost principalele criterii de includere a unei alge verzi în descendența carofitelor (Mattox și Stewart, 1984; Lewis și McCourt, 2004; Leliaert et al., 2012). Din anii 1990 până în prezent, studiile axate pe analizele moleculare ale genomului și transcriptomului cloroplastic și nuclear au reafirmat și rafinat schemele taxonomice și filogenetice anterioare (Timme et al., 2012; Delwiche și Cooper, 2015). Acest lucru a fost completat, în continuare, de analize biochimice, imunologice și bazate pe biologia celulară detaliate ale pereților celulari și ale căilor de biosinteză și de semnalizare a hormonilor din carofite (Popper și Fry, 2003; Popper, 2008; Sørensen et al., 2010, 2011, 2012; Zhang și van Duijn, 2014; Ju et al., 2015; O’Rourke et al., 2015). Carophytele existente prezintă un procent relativ scăzut de diversitate în comparație cu alți taxoni de alge verzi și cuprind 13 familii și 122 de genuri (Becker și Marin, 2009; Leliaert et al., 2012). Opinia filogenetică actuală plasează charophytele în șase clase (figura 1; Delwiche și Cooper, 2015). Clasa bazală, Mesostigmatophyceae, este reprezentată de un singur gen cunoscut, Mesostigma. Această algă este o biflagelată unicelulară cu o formă asimetrică unică (adică, ca o barcă de salvare aplatizată) și este acoperită de straturi de solzi ornamentați (Manton și Ettl, 1965; Becker et al., 1991; Domozych et al., 1991, 1992). Cea de-a doua clasă, Chlorokybophyceae, cuprinde, de asemenea, un singur tip cunoscut, Chlorokybus atmophyticus. Această algă rară formează un pachet sarcinoid de celule înconjurate de un înveliș gros de tip gel (Rogers et al., 1980). Trebuie remarcat faptul că filogeniile alternative plasează Mesostigma și Chlorokybus ca neamuri surori care, împreună, sunt surori cu toate celelalte streptofite (Rodríguez-Ezpeleta et al., 2007). În filogenia celor șase clase, cea de-a treia clasă, Klebsormidiophyceae, este formată din trei genuri care formează filamente simple neramificate (Sluiman et al., 2008), care sunt adesea membre ale „crustelor biologice” care cresc pe suprafețele diferitelor habitate terestre. Mai multe specii din această clasă pot rezista chiar la un stres de desecare semnificativ atunci când fac parte din crustele deșertice (Mikhailyuk et al., 2008, 2014; Holzinger și Karsten, 2013); altele sunt tolerante la stresul de desecare asociat cu mediile cu temperaturi scăzute (Stamenkovic et al., 2014; Herburger și Holzinger, 2015). Aceste prime trei clase constituie carofitele „divergente timpurii”. Linia „divergentă târzie” include, de asemenea, trei clase. Charophyceae, sau stoneworts, se găsesc în mod obișnuit în ecosistemele de apă dulce și posedă talii multicelulari macroscopici care constau în agregate de filamente ramificate alcătuite din celule excepțional de alungite (Lewis și McCourt, 2004). Membrii acestui grup prezintă o reproducere sexuală bazată pe oogamie, care include spermatozoizi mobili și ovule nemotile, ambele produse în gametangii multicelulare. Coleochaetophyceae constă în taxoni care au talii multicelulare formate din talii filamentoși ramificați sau asemănători parenchimului (Graham, 1993; Delwiche et al., 2002). Aceste organisme prezintă, de asemenea, o reproducere sexuală distinctă bazată pe oogamie. Coleochaetophyceae se găsesc, de obicei, la interfața dintre habitatele de apă dulce și cele terestre, adesea ca epifite. În cele din urmă, cel mai mare și mai divers grup de carofite, Zygnematophyceae, este format din unicelule și filamente neramificate (Gontcharov et al., 2003; Guiry, 2013). Caracteristica distinctivă a acestui grup este prezența reproducerii sexuale bazate pe conjugare, care necesită o semnalizare și o aderență celulară complexă (Abe et al., 2016). Algele zygnematofize sunt locuitori comuni ai habitatelor de apă dulce, apărând uneori în înflorituri efemere spectaculoase, precum și în substraturi terestre umede.

FIGURA 1. Charophytes existente. Charophytele existente sunt împărțite în taxoni cu divergență timpurie și tardivă. Taxonii cu divergență timpurie includ Mesostigmatophyceae, Chlorokybophyceae și Klebsormidiophyceae, iar taxonii cu divergență târzie includ Charophyceae, Coleochaetophyceae și Zygnematophyceae. Reprezentanți ai fiecărui grup sunt Mesostigma (Mesostigmatophyceae), Chlorokybus atmophyticus (Chlorokybophyceae), Klebsormidium flaccidum (Klebsormidiophyceae), Chara (Charophyceae), Coleochaete scutata (Coleochaetophyceae) și Micrasterias (Zygnematophyceae).
Carophytes as Model Organisms
Câteva charophytes au fost utilizate pe scară largă ca organisme model în studiul proceselor biologice de bază. Talii lor mici și simpli (de exemplu, în comparație cu plantele terestre) și ușurința în manipularea experimentală sunt doar două dintre atributele lor care le fac organisme model atractive. Dovezi recente au demonstrat, de asemenea, că multe carofite au mai multe caracteristici remarcabil de asemănătoare cu cele ale plantelor terestre, inclusiv prezența căilor de biosinteză pentru mulți regulatori de creștere (Figura 2; Boot et al., 2012; Hori et al., 2014; Wang et al., 2014, 2015; Holzinger și Becker, 2015; Ju et al., 2015) și a mai multor polimeri ai peretelui celular (Popper, 2008; Sørensen et al., 2010, 2011, 2012; Mikkelsen et al., 2014). Aceste două caracteristici au făcut carofitele eficiente în domenii de studiu precum dezvoltarea moleculară a plantelor și fiziologia stresului. Deși multe charophyte au fost utilizate într-o gamă largă de studii biologice, următorii taxoni sunt cei mai notabili pentru utilizarea lor extinsă în mai multe domenii de studiu ca modele:
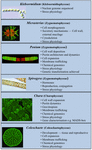
FIGURA 2. Carofite model actuale. Mai multe Charophytes, inclusiv Klebsormidium, Micrasterias, Penium, Spirogyra, Chara și Coleochaete, sunt utilizate ca modele; fiecare dintre ele are atribute specifice care o fac un model deosebit de potrivit pentru procese fiziologice și biochimice specifice.
Desmidele: Modelele simetrice
S-a demonstrat recent că Zygnematophyceae sunt, cel mai probabil, cei mai apropiați strămoși vii ai plantelor terestre (Wodniok et al, 2011; Delwiche și Cooper, 2015). Membrii unicelulari ai grupului incluziv placoderm desmidian au devenit modele importante pentru elucidarea multor principii fundamentale ale biologiei și dezvoltării celulelor vegetale. Dimensiunea mare a celulelor, simetria/forma notabilă, arhitectura distinctă a peretelui celular și sistemele endomembranare/coscheletice elaborate sunt doar câteva dintre caracteristicile care le fac să fie sisteme celulare excelente pentru studierea fiziologiei și dezvoltării celulare. Stabilirea recentă a unor linii transformate stabile și secvențierea în curând a genomurilor/transcriptomelor unor desmide selectate au sporit și mai mult valoarea acestora în studiile botanice. Doi taxoni, Micrasterias și Penium, au apărut ca fiind cei mai bine studiați dintre desmizi.
Micrasterias a fost cel mai popular desmid pentru cercetarea în domeniul biologiei celulare în ultimii 50 de ani datorită caracteristicilor sale unice (Meindl, 1993; Lutz-Meindl, 2016). Celula Micrasterias prezintă o simetrie bilaterală care este adesea evidențiată de o periferie spectaculos disecată (adică, multilobată). Noile semicelule fiice produse prin diviziune celulară nu au această morfologie complexă, ci mai degrabă au o formă sferică. Un program complex de dezvoltare post-citocinetică, care utilizează mai multe seturi de mecanisme subcelulare foarte bine coordonate, generează ulterior fenotipul multilobat în timpul expansiunii semicelulelor fiice. Acest program este, de asemenea, responsabil pentru depunerea de polimeri pentru producerea unui perete celular primar și secundar. Aceste evenimente sunt centrate pe o rețea mare de corpuri Golgi și vezicule asociate care produc un mecanism secretor complex. Acest proces, la rândul său, este direcționat către situsuri specifice ale suprafeței celulare care generează o expansiune celulară multipolară și secreția concomitentă de macromolecule ale peretelui celular și de mucilagii extracelulare (Kim et al., 1996; Lutz-Meindl și Brosch-Salomon, 2000; Oertel et al., 2004; Aichinger și Lutz-Meindl, 2005). Livrarea componentelor secretorii către loci preciși ai suprafeței celulare necesită un sistem citomotil elaborat bazat pe actinomiozină și este controlat de mai multe cascade de transducție a semnalelor (Meindl et al., 1994; Oertel et al., 2003). În plus, morfogeneza celulară este foarte sensibilă la stresul extern (de exemplu, stresul oxidativ, salinitatea, metalele grele) care duce la schimbări majore în diferențierea celulară (Darehshouri et al., 2008; Affenzeller et al., 2009; Andosch et al., 2012). Micrasterias este un organism remarcabil pentru analiza acestor caracteristici subcelulare, deoarece este ușor de întreținut și de manipulat în laborator și se pretează deosebit de bine pentru achiziția de imagini de înaltă rezoluție cu ajutorul microscopiei luminoase și electronice, inclusiv imunocitochimie, microscopie electronică de scanare cu fascicul de ioni focalizat (FIB-SEM) și imagistică prin spectroscopie de pierdere de energie electronică (EELS) (Lutz-Meindl, 2007; Eder și Lutz-Meindl, 2008, 2010; Wanner et al., 2013; Lutz-Meindl et al., 2015). În plus, analizele moleculare inițiale, inclusiv producerea de linii celulare transformate, au sporit și mai mult utilizarea Micrasteriei, în special în dinamica moleculară a procesării peretelui celular (Vannerum et al., 2010, 2011, 2012).
În ultimul deceniu, desmidul, Penium margaritaceum, a devenit, de asemenea, un organism model valoros (Domozych et al., 2009, 2014; Sørensen et al., 2014; Rydahl et al., 2015; Worden et al., 2015). Spre deosebire de Micrasterias, Penium are o formă cilindrică simplă, posedă doar un perete celular primar și depozitează polimeri de perete la doi loci specifici ai suprafeței celulare în timpul expansiunii (Domozych et al., 2011). Această simplitate relativă este foarte atractivă pentru elucidarea principiilor fundamentale ale dezvoltării celulelor vegetale, inclusiv dezvoltarea peretelui celular, expansiunea celulară și dinamica secreției (Domozych et al., 2005, 2014; Ochs et al., 2014). Unul dintre principalele sale atribute este faptul că are polimeri de perete asemănători cu cei prezenți în multe plante terestre (de exemplu, celuloză, pectine, hemiceluloze) și că acești polimeri pot fi marcați cu anticorpi monoclonali. După marcarea celulelor vii, acestea pot fi readuse în cultură, unde pot fi monitorizate evenimentele ulterioare de expansiune celulară și de depunere a peretelui (Domozych et al., 2009; Rydahl et al., 2015). Penium este, de asemenea, ușor de întreținut în laborator, iar ritmul său rapid de creștere în condiții controlate cu precizie îl face un specimen excelent pentru screeninguri de microarray concurente la scară largă ale multor agenți chimici (prin creștere în plăci cu mai multe godeuri) și pentru evaluarea efectelor specifice ale acestora asupra expansiunii/dezvoltării pereților (Worden et al., 2015). Acest lucru ajută în mod semnificativ la dezvăluirea rolului componentelor și proceselor subcelulare specifice în expansiunea/diferențierea. Recent, izolarea unor linii transformate stabile (Sørensen et al., 2014) a sporit și mai mult potențialul acestei alge în viitoarele studii asupra celulelor vegetale.
Chara și Nitella: Cellular Giants for Plant Physiology
Speciile de Chara și Nitella (Charophyceae) au fost folosite de mult timp ca specimene pentru o varietate de investigații biologice, în special cele care se ocupă cu dinamica celulară, expansiunea și fluxul citoplasmatic (Green, 1954; Probine și Preston, 1962). Talii lor macroscopici se disting prin noduri, în care apar ramificațiile și gametangia, și regiuni internodale care constau în celule excepțional de alungite. Celulele internodale prezintă o stratificare clar remarcabilă a componentelor subcelulare, în care cloroplastele orientate elicoidal definesc un cortex staționar care înconjoară o endoplasmă în care are loc cel mai rapid flux citoplasmatic generat de actinomiozină. Citoplasma conține diverse componente endomembranare care participă la rețele dinamice de trafic membranar (de exemplu, endocitoza și exocitoza) care sunt controlate de cascade complexe de transducție a semnalelor (Foissner și Wastenys, 2000, 2012, 2014; Sommer et al., 2015; Foissner et al., 2016). Celulele internodale posedă pereți celulari bogați în celuloză și pectină, homogalacturonan (HG). Turgorul este principala forță motrice pentru expansiune, iar pectina este, cel mai probabil, componenta portantă care controlează expansiunea peretelui. Modularea ciclică, neenzimatică și complexarea concomitentă cu calciu a HG este mecanismul responsabil pentru controlul modulațiilor peretelui care duc la expansiunea celulară (Proseus și Boyer, 2007, 2008, 2012; Boyer, 2016). Unul dintre avantajele tehnice ale Chara și Nitella în cercetarea celulară este că celulele internodale pot fi îndepărtate individual; endoplasma și pereții celulari pot fi „disecate” și apoi folosite ca sisteme celulare pentru manipulări experimentale.
Alte componente ale talilor, cum ar fi rizoizii, au o organizare polară organizată a citoplasmei. Aceste celule conțin statoliți sedimentabili, bogați în minerale, localizați la apex, care funcționează în percepția gravitației (Hodick et al., 1998; Braun, 2002; Braun și Limbach, 2006). Această caracteristică a făcut ca Chara să fie primul carofit din spațiu. În timpul zborurilor parabolice ale rachetei TEXUS, s-a demonstrat că statoliții exercită forțe tensionale asupra filamentelor de actină. Un echilibru de forțe (adică gravitația și forța de contracarare a filamentelor de actină) este responsabil pentru poziționarea corectă a statoliților în rizoid care, la rândul lor, garantează capacitatea de a răspunde la vectorul gravitațional (Braun și Limbach, 2006). Utilizarea pensetelor cu laser și a microscopiei cu centrifugă cu rotație lentă a demonstrat că sedimentarea statoliților nu este suficientă pentru a provoca îndoirea gravitropică a vârfului rizoidului. Mai degrabă, statolitele bogate în minerale trebuie să se așeze pe regiuni specifice ale membranei plasmatice pentru ca efectele morfologice gravitropice să aibă loc (Braun, 2002). Chara a fost, de asemenea, utilizată pentru a studia alte fenomene, inclusiv electrofiziologia și rolul și mișcarea hormonilor (Belby, 2016).
Coleochaete for Developmental Studies and Pattern Development
Formarea unui thallus (adică a unui țesut, organ) multicelular al unei plante necesită planuri de diviziune celulară precise, reglementate spațial, care sunt controlate de mai multe seturi de gene exprimate în puncte specifice în timpul dezvoltării (Besson și Dumais, 2011; Umen, 2014). Direcționată atât în spațiu tridimensional, cât și în timp, morfogeneza talusului se modulează, de asemenea, ca răspuns la factorii externi de stres. La plante, aceste programe de dezvoltare sunt și mai complexe din cauza prezenței pereților celulari care nu permit migrarea celulelor sau flexibilitatea țesuturilor. Elucidarea evenimentelor specifice care sunt esențiale pentru manifestarea formei și dimensiunii talusului este adesea dificil de descifrat la plantele terestre, unde rezoluția comportamentelor specifice ale celulelor este adesea slab rezolvată atunci când sunt încorporate în țesuturi/organe foarte complexe și expansive. Utilizarea organismelor cu talusuri mai simple s-a dovedit a fi avantajoasă pentru înțelegerea fenomenelor de bază ale dezvoltării, iar genul Coleochaete de carofite este un astfel de exemplu. Anumite specii (de exemplu, C. orbicularis, C. scutatum) produc un thallus discoid parenchimatos care crește în exterior printr-o combinație de diviziuni celulare anti- sau peri-clinale și de expansiune ulterioară a celulelor cele mai exterioare (Brown et al., 1994; Cook, 2004). Planul de diviziune al oricărei celule urmează reguli simple care se bazează pe dimensiunea și forma celulei și pe poziția geografică în talus (Dupuy et al., 2010; Besson și Dumais, 2011). Această caracteristică permite construirea de modele matematice care pot fi apoi utilizate pentru a interpreta morfogeneza multicelulară (Domozych și Domozych, 2014; Umen, 2014) și oferă o perspectivă critică asupra evoluției, biomecanicii și fiziologiei organelor și organismelor întregi. Mai mult, unele specii de Coleochaete au fost studiate pe larg pentru a determina modulațiile structurale și funcționale care apar la un thallus în condiții de desecare (Graham et al., 2012), adică caracteristici esențiale pentru înțelegerea evoluției timpurii a plantelor terestre și a dinamicii creșterii plantelor în timpul secetei.
Insights from Genomic and Gene Studies
În mod similar cu multe alte tehnologii, instrumentele moleculare au capacitatea de a transmite noi informații importante pentru multe domenii de cercetare, inclusiv fiziologia și dezvoltarea. De exemplu, membri ai familiei de gene MADS-box au fost izolați și caracterizați de la trei carofite, Chara globularis, Coleochaete scutata și Closterium peracerosum-strigosum-littorale (Tanabe et al., 2005). Modelul de expresie al genelor MADS-box la carofite sugerează că acestea joacă un rol în dezvoltarea și reproducerea haploidă (Tanabe et al., 2005). Se crede că acestea au fost recrutate în generația diploidă, iar rolurile lor în dezvoltare s-au diversificat în continuare, în timpul evoluției plantelor terestre (Tanabe et al., 2005). Până în prezent, Klebsormidium flaccidum este singurul carofit pentru care este disponibil un genom nuclear aproape complet (Hori et al., 2014); deși genomurile cloroplastului și mitocondriale au fost secvențiate pentru reprezentanți ai fiecăreia dintre cele șase clase de carofite și sunt disponibile EST-uri (expressed sequence tags) pentru unele carofite (Delwiche, 2016). Genomul Klebsormidium a dezvăluit deja prezența genelor pentru sinteza mai multor hormoni vegetali și intermediari de semnalizare, mecanisme de protecție împotriva intensității luminoase ridicate (Hori et al., 2014) și prezența factorilor de transcripție WRKY din grupul IIb, despre care s-a crezut anterior că au apărut pentru prima dată la mușchi (Rinerson et al., 2015). Mai multe proiecte de secvențiere a genomului carofitelor sunt în curs de desfășurare (Delwiche, 2016) și vor permite efectuarea unor analize comparative mai detaliate, incluzând un număr mai mare de carofite și, potențial, vor oferi noi informații despre relațiile evolutive dintre specii și gene. Poziția unică a carofitelor ca fiind cel mai timpuriu grup divergent de streptofite face din ele un instrument deosebit de puternic pentru studiile de evoluție-dezvoltare și tehnologiile emergente, cum ar fi realizarea recentă a transformării stabile la Penium (Sørensen et al, 2014) vor permite explorarea funcției unor gene specifice.
Future Uses of Charophytes as Model Organisms
Un următor pas esențial în cercetarea carofitelor este analiza genomurilor taxonilor incluși și studiile comparative cu cele ale plantelor terestre. Nu numai că această lucrare va oferi informații esențiale despre mecanismele de adaptare pe care carofitele le-au evoluat atunci când au apărut pe habitatele terestre și le-au colonizat cu succes, dar va perfecționa și va extinde și mai mult utilizarea organismelor model actuale în cercetarea fundamentală a plantelor. Deși studiile moleculare vor sprijini multe domenii de cercetare, două domenii care vor beneficia imediat vor fi biologia peretelui celular și dinamica regulatorilor de creștere în creștere și dezvoltare. În primul caz, carofitele ar putea oferi clarificări cu privire la multe fenomene de bază, dar slab rezolvate, inclusiv biosinteza pectinei și a proteinelor din peretele celular, secreția controlată, depunerea și modularea post-secretorie a polimerilor din pereți, precum și rețelele specifice și interactive de trafic membranar din celulele vegetale. În acest din urmă domeniu, modelele de carofite ar ajuta la descifrarea semnalizării moleculare, a biologiei celulare și a dinamicii de dezvoltare a creșterii și dezvoltării asociate cu hormoni precum etilena și auxina. Studiile ecofiziologice recente cu carofite oferă, de asemenea, o potențială bogăție de date critice în determinarea modului în care celulele vegetale se adaptează la stres, inclusiv la desecare și la toleranța la sare (Holzinger și Karsten, 2013; Pichrtová et al., 2014; Herburger et al., 2015; Holzinger și Becker, 2015; Holzinger și Pichrtová, 2016; Kondo et al., 2016). Viitorul cercetării carofitelor este, într-adevăr, foarte luminos și va deveni, în consecință, o binefacere pentru întreaga biologie vegetală.
Contribuții ale autorilor
Toți autorii enumerați, au avut o contribuție substanțială, directă și intelectuală la lucrare și au aprobat-o pentru publicare.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoaștere
Această lucrare a fost susținută de granturile de colaborare NSF (SUA) MCB 1517345 și 1517546.
Belby, M. J. (2016). Sistem experimental de characee la scară multiplă: de la electrofiziologia transportatorilor de membrană la conectivitatea celulă-la-celulă, fluxul citoplasmatic și metabolismul auxinei. Front. Plant Sci. 7:1052. doi: 10.3389/fpls.2016.01052
CrossRef Full Text | Google Scholar
Braun, M. (2002). Percepția gravitației necesită statoliți așezați pe zone specifice ale membranei plasmatice în rizoidele și protonematele characeelor. Protoplasma 219, 150-159. doi: 10.1007/s007090200016
CrossRef Full Text | Google Scholar
Braun, M., și Limbach, C. (2006). Rhizoidele și protonematele algelor characee: celule model pentru cercetări privind creșterea polarizată și detectarea gravitației plantelor. Protoplasma 229, 133-142. doi: 10.1007/s00709-006-0208-9
CrossRef Full Text | Google Scholar
Graham, L. E. (1993). Originea plantelor terestre. New York, NY: John Wiley & Sons, 287.
Google Scholar
Herburger, K., și Holzinger, A. (2015). Localizarea și cuantificarea calosei în algele verzi streptofite Zygnema și Klebsormidium: corelație cu toleranța la desecare. Plant Cell Physiol. 56, 2259-2270. doi: 10.1093/pcp/pcv139
CrossRef Full Text | Google Scholar
Lemieux, C., Otis, C., și Turmel, M. (2016). Analizele comparative ale genomului cloroplastului din algele verzi streptofite descoperă modificări structurale majore în Klebsormidiophyceae, Coleochaetophyceae și Zygnematophyceae. Front. Plant Sci. 7:697. doi: 10.3389/fpls.2016.00697
CrossRef Full Text | Google Scholar
Lutz-Meindl, U. (2007). Utilizarea microscopiei electronice cu transmisie cu filtrare de energie pentru generarea de imagini și analiza elementelor în organismele vegetale. Micron 38, 181-196. doi: 10.1016/j.micron.2006.03.017
CrossRef Full Text | Google Scholar
Meindl, U. (1993). Celulele Micrasterias ca sistem model pentru cercetarea morfogenezei. Microbiol. Rev. 57, 415-433.
Google Scholar
Pichrtová, M., Kulichova, J., și Holzinger, A. (2014). Limitarea azotului și uscarea lentă induc toleranța la desecare în algele verzi conjugate (Zygnematophyceae, Streptophyta) din habitatele polare. PLoS ONE 9:e113137. doi: 10.1371/journal.pone.0113137
CrossRef Full Text | Google Scholar
Pickett-Heaps, J. D., and Marchant, H. J. J. (1972). The phylogeny of green algae: a new proposal (Filogenia algelor verzi: o nouă propunere). Cytobios 6, 255-264.
Google Scholar
Stamenkovic, M., Woelken, E., și Hanelt, D. (2014). Ultrastructura tulpinilor de Cosmarium (Zygnematophyceae, Streptiophyta) colectate din diferite locații geografice arată diferențe specifice speciilor atât la temperaturi optime, cât și la temperaturi de stres. Protoplasma 251, 1491-1509.
Google Scholar
.