Chemolithotrophie
Chemolithotrophie ist die Oxidation von anorganischen Chemikalien zur Energiegewinnung. Der Prozess kann die oxidative Phosphorylierung nutzen, genau wie die aerobe und anaerobe Atmung, aber jetzt ist die Substanz, die oxidiert wird (der Elektronendonator) eine anorganische Verbindung. Die Elektronen werden an Ladungsträger innerhalb der Elektronentransportkette weitergegeben und erzeugen eine Protonenmotivationskraft, die mit Hilfe der ATP-Synthase zur Erzeugung von ATP genutzt wird.
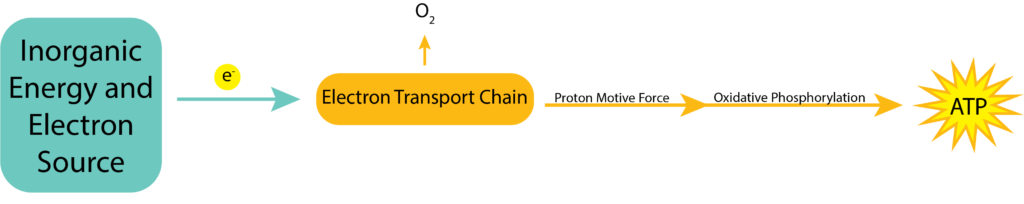
Elektronendonatoren
Chemolithotrophe Organismen verwenden eine Vielzahl anorganischer Verbindungen als Elektronendonatoren, wobei Wasserstoffgas, Schwefelverbindungen (wie Sulfid und Schwefel), Stickstoffverbindungen (wie Ammonium und Nitrit) und Eisen(II) am häufigsten vorkommen.
- Wasserstoffoxidierer – diese Organismen oxidieren Wasserstoffgas (H2) mit Hilfe eines Hydrogenase-Enzyms. Es gibt sowohl aerobe als auch anaerobe Wasserstoffoxidierer, wobei die aeroben Organismen schließlich Sauerstoff zu Wasser reduzieren.
- Schwefeloxidierer – als Gruppe sind diese Organismen in der Lage, eine Vielzahl von reduzierten und teilweise reduzierten Schwefelverbindungen wie Schwefelwasserstoff (H2S), elementaren Schwefel (S0), Thiosulfat (S2O32-) und Sulfit (SO32-) zu oxidieren. Sulfat (SO42-) ist häufig ein Nebenprodukt der Oxidation. Oft erfolgt die Oxidation schrittweise mit Hilfe des Enzyms Sulfit-Oxidase.
- Stickstoffoxidierer – die Oxidation von Ammoniak (NH3) erfolgt in einem zweistufigen Prozess durch nitrifizierende Mikroben, wobei eine Gruppe Ammoniak zu Nitrit (NO2-) und die zweite Gruppe das Nitrit zu Nitrat (NO3-) oxidiert. Der gesamte Prozess ist als Nitrifikation bekannt und wird von kleinen Gruppen aerober Bakterien und Archaeen durchgeführt, die oft gemeinsam im Boden oder in Wassersystemen leben.
- Eisenoxidierer – diese Organismen oxidieren Eisen (Fe2+) zu Eisen(III) (Fe3+). Da Fe2+ ein so positives Standard-Reduktionspotential hat, ist die Bioenergetik nicht besonders günstig, selbst wenn Sauerstoff als letzter Elektronenakzeptor verwendet wird. Die Situation wird für diese Organismen dadurch erschwert, dass Fe2+ in Gegenwart von Sauerstoff spontan zu Fe3+ oxidiert; die Organismen müssen es für ihre eigenen Zwecke nutzen, bevor das geschieht.
Elektronenakzeptoren
Chemolithotrophie kann aerob oder anaerob erfolgen. Wie bei beiden Atmungsarten ist der beste Elektronenakzeptor Sauerstoff, um den größten Abstand zwischen Elektronendonator und Elektronenakzeptor zu schaffen. Die Verwendung eines Nicht-Sauerstoff-Akzeptors ermöglicht den chemolithotrophen Organismen eine größere Vielfalt und die Fähigkeit, in einer größeren Vielfalt von Umgebungen zu leben, auch wenn sie dafür Abstriche bei der Energieproduktion machen müssen.
Menge des erzeugten ATP
So wie sowohl die Elektronendonatoren als auch die -akzeptoren bei dieser Gruppe von Organismen stark variieren können, wird auch die Menge des erzeugten ATP für ihre Bemühungen stark variieren. Sie werden nicht so viel ATP erzeugen wie ein Organismus, der die aerobe Atmung nutzt, da das größte ΔE0′ mit Glukose als Elektronendonor und Sauerstoff als Elektronenakzeptor erzielt wird. Wie viel weniger als 32 ATP-Moleküle es sind, hängt jedoch in hohem Maße von dem tatsächlich verwendeten Donor und Akzeptor ab. Je geringer der Abstand zwischen den beiden, desto weniger ATP wird gebildet.
Chemolithoautotrophe vs. Chemolithoheterotrophe
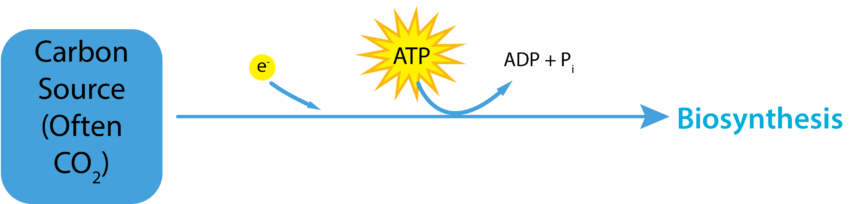
Die meisten Chemolithotrophen sind Autotrophe (Chemolithoautotrophe), bei denen sie atmosphärisches Kohlendioxid fixieren, um die von ihnen benötigten organischen Verbindungen aufzubauen. Diese Organismen benötigen sowohl ATP als auch Reduktionskraft (d. h. NADH/NADPH), um das oxidierte Molekül CO2 schließlich in eine stark reduzierte organische Verbindung wie Glukose umzuwandeln. Wenn eine chemolithoautotrophe Pflanze einen Elektronendonator mit einem höheren Redoxpotential als NAD+/NADP verwendet, muss sie den umgekehrten Elektronenfluss nutzen, um die Elektronen im Elektronenturm wieder nach oben zu befördern. Dies ist energetisch ungünstig für die Zelle, da sie Energie aus dem Protonenantrieb verbraucht, um die Elektronen in umgekehrter Richtung durch die ETC zurückzutreiben.
Einige Mikroben sind chemolithoheterotroph, sie verwenden eine anorganische Chemikalie für ihren Energie- und Elektronenbedarf, sind aber auf organische Chemikalien in der Umgebung angewiesen, um ihren Kohlenstoffbedarf zu decken. Diese Organismen werden auch als mixotroph bezeichnet, da sie für ihr Wachstum und ihre Fortpflanzung sowohl anorganische als auch organische chemische Verbindungen benötigen.
Stickstoff-Stoffwechsel
Der Stickstoffkreislauf beschreibt die verschiedenen Wege, auf denen Stickstoff, ein lebenswichtiges Element, von Organismen für verschiedene Zwecke genutzt und umgewandelt wird. Ein Großteil der chemischen Umwandlungen wird von Mikroben im Rahmen ihres Stoffwechsels durchgeführt, wobei sie anderen Organismen einen wertvollen Dienst erweisen, indem sie sie mit einer alternativen chemischen Form des Elements versorgen.
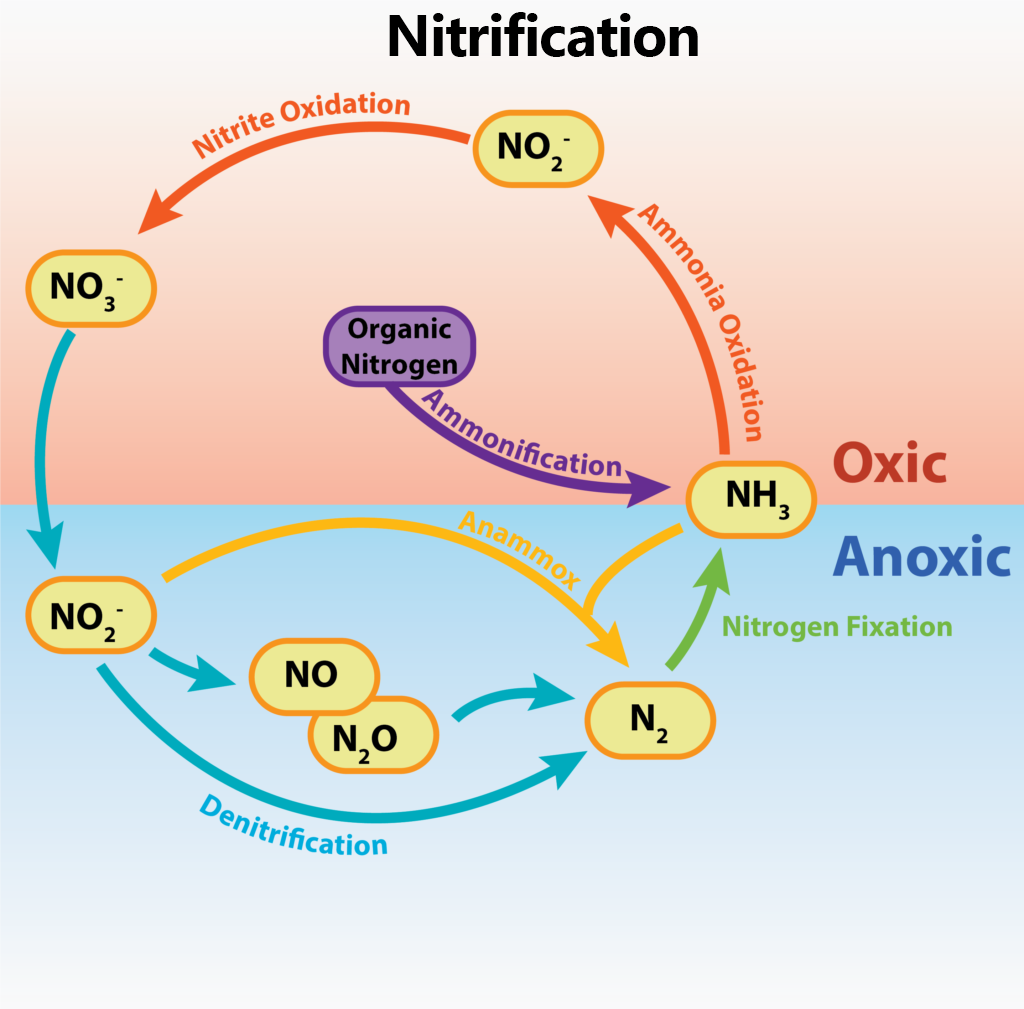
Stickstofffixierung
Die Stickstofffixierung beschreibt die Umwandlung des relativ reaktionsträgen Gases Distickstoff (N2) in Ammoniak (NH3), eine für die meisten Lebensformen wesentlich besser nutzbare Form von Stickstoff. Dieser Prozess wird von diazotrophen Organismen durchgeführt, einer begrenzten Anzahl von Bakterien und Archaeen, die aufgrund ihrer Fähigkeiten ohne eine externe Quelle von fixiertem Stickstoff wachsen können. Die Stickstofffixierung ist ein wesentlicher Prozess für die Organismen der Erde, da Stickstoff ein notwendiger Bestandteil verschiedener organischer Moleküle, wie Aminosäuren und Nukleotide, ist. Pflanzen, Tiere und andere Organismen sind auf Bakterien und Archaeen angewiesen, um Stickstoff in fixierter Form bereitzustellen, da kein Eukaryote bekannt ist, der Stickstoff fixieren kann.
Die Stickstofffixierung ist ein extrem energie- und elektronenintensiver Prozess, um die Dreifachbindung in N2 zu brechen und es zu NH3 zu reduzieren. Dazu ist ein bestimmtes Enzym, die Nitrogenase, erforderlich, das durch O2 inaktiviert wird. Daher muss die Stickstofffixierung in einer anaeroben Umgebung stattfinden. Aerobe stickstofffixierende Organismen müssen besondere Bedingungen oder Vorkehrungen treffen, um ihr Enzym zu schützen. Stickstofffixierende Organismen können entweder unabhängig voneinander existieren oder sich mit einem pflanzlichen Wirt zusammentun:
- Symbiotische stickstofffixierende Organismen: Diese Bakterien gehen eine Partnerschaft mit einer Pflanze ein, um ihr ein geeignetes Umfeld für das Funktionieren ihres Stickstoffase-Enzyms zu bieten. Die Bakterien leben im Gewebe der Pflanze, oft in Wurzelknöllchen, binden Stickstoff und teilen die Ergebnisse. Die Pflanze stellt sowohl den Ort für die Stickstofffixierung als auch zusätzliche Nährstoffe zur Verfügung, um den energieaufwändigen Prozess der Stickstofffixierung zu unterstützen. Es hat sich gezeigt, dass die Bakterien und der Wirt chemische Erkennungssignale austauschen, die diese Beziehung erleichtern. Eines der bekanntesten Bakterien dieser Kategorie ist Rhizobium, das sich mit Pflanzen aus der Familie der Hülsenfrüchte (Klee, Soja, Luzerne usw.) zusammentut.
- Freilebende stickstofffixierende Organismen: Diese Organismen, sowohl Bakterien als auch Archaeen, fixieren Stickstoff für ihren eigenen Bedarf, der schließlich weitergegeben wird, wenn der Organismus stirbt oder aufgenommen wird. Freilebende stickstofffixierende Organismen, die anaerob wachsen, müssen sich nicht um spezielle Anpassungen für ihr Nitrogenase-Enzym kümmern. Aerobe Organismen müssen Anpassungen vornehmen. Cyanobakterien, ein mehrzelliges Bakterium, bilden spezialisierte Zellen, die als Heterocysten bekannt sind und in denen die Stickstofffixierung stattfindet. Da Cyanobakterien im Rahmen ihrer Photosynthese Sauerstoff produzieren, entsteht in der Heterozyste eine sauerstofffreie Version, so dass die Nitrogenase aktiv bleiben kann. Die Heterozysten teilen den fixierten Stickstoff mit den umliegenden Zellen, während die umliegenden Zellen die Heterozysten mit zusätzlichen Nährstoffen versorgen.
Assimilation
Assimilation ist ein reduktiver Prozess, bei dem eine anorganische Form von Stickstoff zu organischen Stickstoffverbindungen wie Aminosäuren und Nukleotiden reduziert wird, was zelluläres Wachstum und Reproduktion ermöglicht. Es wird nur die von der Zelle benötigte Menge reduziert. Ammoniak-Assimilation liegt vor, wenn das bei der Stickstofffixierung gebildete Ammoniak (NH3)/Ammonium-Ion (NH4+) in den zellulären Stickstoff eingebaut wird. Die assimilative Nitratreduktion ist eine Reduktion von Nitrat zu zellulärem Stickstoff in einem mehrstufigen Prozess, bei dem Nitrat zu Nitrit, dann zu Ammoniak und schließlich zu organischem Stickstoff reduziert wird.
Nitrifikation
Wie oben erwähnt, wird die Nitrifikation von chemolithotrophen Organismen durchgeführt, die eine reduzierte oder teilweise reduzierte Form von Stickstoff als Elektronendonor zur Energiegewinnung nutzen. ATP wird durch den Prozess der oxidativen Phosphorylierung unter Verwendung von ETC, PMF und ATP-Synthase gewonnen.
Denitrifikation
Denitrifikation bezieht sich auf die Reduktion von NO3- zu gasförmigen Stickstoffverbindungen, wie N2. Denitrifizierende Mikroben führen eine anaerobe Atmung durch, wobei sie NO3- als alternativen Endelektronenakzeptor zu O2 verwenden. Dabei handelt es sich um eine Art dissimilatorische Nitratreduktion, bei der das Nitrat zur Energieerhaltung und nicht zur Herstellung organischer Verbindungen reduziert wird. Dabei entstehen große Mengen an überschüssigen Nebenprodukten, die dazu führen, dass Stickstoff aus der lokalen Umgebung an die Atmosphäre abgegeben wird.
Anammox
Anammox oder anaerobe Ammoniakoxidation wird von relativ neu entdeckten Meeresbakterien durchgeführt, die Stickstoffverbindungen sowohl als Elektronenakzeptor als auch als Elektronendonator nutzen. Ammoniak wird anaerob als Elektronendonor oxidiert, während Nitrit als Elektronenakzeptor genutzt wird, wobei als Nebenprodukt Distickstoffgas entsteht. Die Reaktionen laufen innerhalb des Anammoxosoms ab, einer spezialisierten Zytoplasmastruktur, die 50-70 % des gesamten Zellvolumens ausmacht. Genau wie bei der Denitrifikation wird bei der Anammox-Reaktion gebundener Stickstoff aus der lokalen Umgebung entfernt und an die Atmosphäre abgegeben.
Schlüsselwörter
Chemolithotrophie, Wasserstoffoxidierer, Hydrogenase, Schwefeloxidierer, Sulfitoxidierer, Stickstoffoxidierer, Nitrifikation, Eisenoxidierer, chemolithoautotroph, umgekehrter Elektronenfluss, chemolithoheterotroph, mixotroph, Stickstofffixierung, diazotroph, Nitrogenase, symbiotische stickstofffixierende Organismen, Rhizobium, Leguminose, freilebende stickstofffixierende Organismen, Cyanobakterien, Heterozyste, Assimilation, Ammoniakassimilation, assimilative Nitratreduktion, Denitrifikation, dissimilatorische Nitratreduktion, Anammox, anaerobe Ammoniakoxidation, Anammoxosom.
Studienfragen
- Was ist Chemolithotrophie?
- Was sind die häufigsten Elektronendonatoren und -akzeptoren bei Chemolithotrophen? Wie ist die Menge des produzierten ATP im Vergleich zu chemoorganotrophen Organismen?
- Wie unterscheiden sich chemolithoautotrophe und chemolithoheterotrophe Organismen? Was ist der umgekehrte Elektronenfluss und wie/warum wird er von einigen Chemolithoautotrophen genutzt?
- Welche Rolle spielen Bakterien/Archeen im Stickstoffkreislauf? Wie werden verschiedene Stickstoffverbindungen in ihrem Stoffwechsel verwendet?
- Was ist für die Stickstofffixierung erforderlich? Wie unterscheiden sich frei lebende Stickstofffixierer und pflanzenassoziierte Stickstofffixierer? Wie schützen Rhizobium und Cyanobakterien ihre Nitrogenase vor Sauerstoff?
- Was sind die verschiedenen Mechanismen des Stickstoffmetabolismus? Welche Umwandlung findet bei jedem statt? Was ist der Zweck jedes einzelnen Mechanismus und wie hängt er mit dem Stoffwechsel des Organismus zusammen?