Introduktion
Tidligere forskning viser, at mennesker kan bruge et stikord til mentalt at forberede sig på en kognitiv udfordring. Hvis deltagerne instrueres i at forberede sig mentalt i det øjeblik, hvor et cue præsenteres, er dette blevet defineret som fasisk årvågenhed . Evnen til at påvirke vores niveau af årvågenhed kunne være vigtig, især hvis årvågenhed også kommer med en omkostning. Hvis kravet til årvågenhed er for højt, kan det potentielt føre til træthed (Härmä et al., 2008) eller stressrelaterede problemer. I denne undersøgelse har vi til formål at undersøge, om og hvordan sunde voksne afstemmer deres niveau af årvågenhed til den forventede vanskelighed af en kommende kognitiv opgave.
Hvis et cue præsenteres lige før en opgavestimulus, er folk generelt i stand til at reagere hurtigere i enkle motoriske responsopgaver (Fan et al., 2002; Macleod et al., 2010; Weinbach og Henik, 2011) og opfatter degraderede visuelle stimuli mere nøjagtigt (Kusnir et al., 2011). Billeddannelsesundersøgelser har vist, at fasisk årvågenhed er forbundet med øget aktivering i frontale, parietale, thalamiske (Shulman et al., 1999; Fan et al., 2005; Yanaka et al., 2010), temporo-occipitale (Thiel et al., 2004) og visuelle hjerneområder (Bartolucci og Smith, 2011). Disse billeddannelsesundersøgelser har givet værdifulde oplysninger om repræsentationen af fasisk alarmberedskab i hjernen. Imidlertid er fasisk årvågenhed generelt blevet undersøgt som et on/off-fænomen, eller analysen er blevet begrænset til den visuelle cortex. Derfor er det uklart, om årvågenhed også afspejler den forventede sværhedsgrad af den kognitive udfordring i hele hjernenetværk.
Dertil kommer, at tidligere undersøgelser har vist, at deaktivering af hjerneområder, der er en del af default mode-netværket (DMN), kan lette udførelsen af kognitive opgaver. For det første viser undersøgelser af opgaveudførelse, at et stigende niveau af opgavens sværhedsgrad er forbundet med stigende deaktivering (McKiernan et al., 2003, 2006; Jansma et al., 2007; Singh og Fawcett, 2008; Pyka et al., 2009; Hedden et al., 2012; Čeko et al., 2015). For det andet har Jansma et al. (2007) vist, at den mediale præfrontale del af DMN kun afstemmer til opgavens vanskelighed, som deltageren kunne forudse, men blev ikke påvirket af den faktiske vanskelighed af hver stimulus, som deltageren ikke kunne forudse. Endelig fandt Weissman et al. (2006), at deaktivering i DMN lige før stimulus var svagere, hvis forsøgspersoner reagerede relativt langsomt. Måske reagerede disse forsøgspersoner langsommere, fordi de midlertidigt var mindre opmærksomme.
For vores undersøgelse designede vi en cued visuel opfattelsesopgave med en let og vanskelig betingelse. Cues gav information om sværhedsgraden af den kommende stimulus, men gav ikke nogen information, der kunne lette selve opgaven. I halvdelen af forsøgene blev cue’et ikke efterfulgt af en stimulus. Kun disse forsøg blev analyseret for hjerneaktivitet, så vi helt kunne isolere aktivering forbundet med cue’et fra aktivering forbundet med opgavens udførelse. Vi antager, at deltagerne afstemmer deres aktivitet til den forventede vanskelighed af opgaven.
Materialer og metoder
Deltagere
Deltagerne blev rekrutteret via online-annoncering. Deltagerne blev udelukket, hvis de rapporterede en historie med betydelige neurologiske eller psykiatriske lidelser eller kontraindikationer for MR-scanning (magnetisk resonansbilleddannelse (MRI) (metalgenstande i eller omkring kroppen, klaustrofobi eller graviditet). Denne undersøgelse blev gennemført i overensstemmelse med anbefalingerne i loven om medicinsk forskning med deltagelse af mennesker (WMO), Medicinsk forskningsetisk komité for medicinsk forskning i Brabant. Protokollen blev godkendt af den etiske komité for medicinsk forskning (protokolnummer: NL51147.028.14). Alle forsøgspersoner gav skriftligt informeret samtykke i overensstemmelse med Helsinki-erklæringen.
Togtyve sunde højrehåndede frivillige deltog i undersøgelsen. Data fra to deltagere blev udelukket på grund af scannerartefakter. Resultater rapporteres for de resterende 20 deltagere .
Task Design
Vi designede en opgave (med begivenhedsrelateret design), der gjorde det muligt for os at undersøge forventningseffekter relateret til den forventede vanskelighed af en opgave, uden forvirrende effekter af udførelsen af opgaven.
Task-stimuli bestod af ni pile i et tre gange tre layout på en sort skærm (Figur 1). En “let stimulus” indeholdt otte pile i den rigtige retning og en pil i den modsatte retning. En “vanskelig stimulus” indeholdt fem pile i den rigtige retning og fire pile i den modsatte retning (figur 1). Deltagerne blev instrueret om at trykke på en knap med den hånd, der svarer til den retning, som flertallet af pilene havde. Nemme og svære stimuli blev præsenteret i en tilfældig rækkefølge. Baseline-stimuli bestod af en stationær sort skærm med teksten: “du har en 30 s pause” på hollandsk og med hvide bogstaver. Stimuli blev præsenteret af “præsentations”-software.
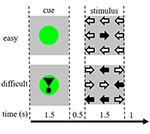
Figur 1. Repræsentation af de visuelle stimuli. Hvert forsøg var på 4,5 s. I halvdelen af forsøgene blev cue’et ikke efterfulgt af en stimulus. Disse forsøg blev anvendt til analysen af funktionel magnetisk resonansbilleddannelse (fMRI).
Hvert forsøg startede med et cue, der angav sværhedsgraden af den efterfølgende præsenterede stimulus. En grøn prik indikerede en “let stimulus”, en grøn prik med et udråbstegn indikerede en “svær stimulus” (Figur 1).
I halvdelen af forsøgene blev cueet ikke efterfulgt af en stimulus, i den anden halvdel blev stimulus præsenteret 500 ms efter at cueet forsvandt. Dette resulterede i fire betingelser: (1) et cue for en let stimulus, ikke efterfulgt af en stimulus (“CueE”); (2) et cue for en vanskelig stimulus, ikke efterfulgt af en stimulus (“CueD”); (3) et cue for en let stimulus, efterfulgt af en let stimulus (“StimE”); og (4) et cue for en vanskelig stimulus, efterfulgt af en vanskelig stimulus (“StimD”). Alle betingelser blev kontrasteret til baseline. Funktionelle MRI-resultater (fMRI) var baseret på CueE og CueD. Præstationsresultater var baseret på StimE og StimD. CueE og CueD blev præsenteret i en pseudo-randomiseret rækkefølge ved hjælp af M-sekvenser for at maksimere generel lineær model (GLM) regressoruafhængighed mellem betingelser og optimere designets effektivitet (Buracas og Boynton, 2002). Deltagerne kunne ikke forudsige, om et cue ville blive fulgt af en stimulus eller ej.
Den varighed af hvert forsøg var 4,500 ms. Cues blev præsenteret ved t = 0 i 1.500 ms, og stimuli blev præsenteret ved t = 2.000, også i 1.500 ms (figur 1). Opgaven blev udført i to omgange af 68 forsøg. Baseline bestod af tre 30 s hvileperioder, før, efter og mellem de to kørsler. Den samlede varighed af opgaven var 11,7 min. Dette var den anden opgave i sessionen og startede, efter at deltagerne havde ligget ca. 10 min i scanneren.
Deltagerne blev instrueret om at forberede sig på en let eller vanskelig stimulus baseret på oplysningerne i cue’et (Figur 1) og at angive så præcist og hurtigt som muligt retningen af flertallet af pilene i stimulus. Deltagerne øvede opgaven uden for scanneren efter en standardøvelsesprotokol på 3 min. Denne protokol blev gentaget, hvis præstationen var utilstrækkelig (under 70 % nøjagtighed for de vanskelige stimuli).
Et spejl, der var fastgjort til hovedspolen, gjorde det muligt for deltagerne at se en gennemsigtig projektionsskærm, der var placeret bag hovedet. En videoprojektor inde i scannerrummet projicerede opgavestimuliene på skærmen. Der blev anvendt to pneumatiske trykknapbokse med lufttryk til at registrere reaktioner. Deltagerne brugte venstre tommelfinger eller pegefinger til at svare “venstre” og højre til at svare “højre”.”
Billedindsamling
Scanninger blev udført på en 3T Philips Achieva-scanner (Philips Medical Systems, Best, Holland) ved hjælp af en 32-kanals SENSE-hovedspole. Et 3D T1-vægtet strukturelt billede blev erhvervet med henblik på anatomisk registrering . fMRI-billeder blev opnået ved hjælp af en 3D PRESTO-pulssekvens . Seks dummy scanninger blev erhvervet og derefter kasseret af scanneren.
Billedforbehandling
fMRI-data blev forbehandlet ved hjælp af statistisk parametrisk kortlægning (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, London, UK2). Scanninger fra en session blev justeret til den første scanning for at korrigere for emnebevægelse ved hjælp af en mindste kvadraters tilgang, en seks parameter (rigid body) rumlig transformation og en 2. grads B-spline-estimation. Scanningerne blev samregistreret til T1 ved hjælp af en rigid kropsmodel. Parametrene blev estimeret ved hjælp af den normaliserede gensidige informationsfunktion. Billederne blev omskåret ved hjælp af en B-spline af 4. grad. T1 blev rumligt normaliseret til standard MNI-rum ved hjælp af en meget let bias-regularisering (0,0001) og en 4. grads B-Spline. De resulterende parametre blev anvendt på alle funktionelle scanninger for at minimere anatomiske forskelle og dermed muliggøre gruppeanalyse. Endelig blev alle scanninger rumligt udglattet med et 3D Gaussisk filter (fuld bredde ved halvt maksimum: 8 mm) for yderligere at minimere effekten af funktionelle anatomiske forskelle.
Individuel fMRI-analyse
Eventuel GLM-regressionsanalyse blev udført for hver voxel for at generere individuelle aktiveringskort ved hjælp af en masse-univariat tilgang med en global tilnærmet AR(1) autokorrelationsmodel og et højpasfilter (128 s cut-off). Baseline blev ikke eksplicit modelleret. Den grundlæggende funktion var en kanonisk HRF uden derivater. Der blev anvendt en maskeringstærskel på 0,8. Der blev anvendt særskilte regressorer for hver betingelse (CueE, 17 tidspunkter; CueD, 16 tidspunkter; StimE, 17 tidspunkter; StimD, 18 tidspunkter; og en uhensigtsmæssig regressor for blanke perioder i opgaven i forbindelse med m-sekvensen, 68 tidspunkter). Ingen tidspunkter blev udelukket fra analysen. Betakort blev transformeret for at afspejle den faktiske procentvise signalændring i hver voxel. Vi præsenterer kun resultaterne for CueE- og CueD-regressorer, der afspejler signalændringer for cues uden en stimulus. Beta og statistiske t-maps blev kontrolleret visuelt for større artefakter.
Gruppe fMRI-analyse
En fMRI-analyse på andet niveau blev udført med lineær regression ved hver voxel til visualisering og udvælgelse af region af interesse (ROI). Der blev udført en ROI-analyse med GNU data language (GDL3) ved hjælp af individuelle emneprocentvise signalændringskort, der blev genereret af GLM-analysen. Voxellerne inden for en ROI eller et netværk blev gennemsnitliggjort for at beregne signalforskellen mellem CueE, CueD og baseline.
ROI-valg
Supratentorielle lokale maksima og minima blev bestemt for både CueE og CueD ved SPM med en ukorrigeret tærskel (p < 0,001; Tabel 1). Kubiske ROI’er af foruddefineret størrelse og form blev placeret over de lokale maksima for CueE-baseline- og CueD-baseline-kontrasten separat inden for et foruddefineret raster (Jansma og Rutten, 2017; Tabel 2). ROI’er blev inkluderet i analysen, hvis de indeholdt et signifikant lokalt maksimum for CueE eller CueD. Ved at definere ROI’er på lokale maksima for begge betingelser separat forhindrer vi bias til enten CueE eller CueD. Desuden reducerer den foruddefinerede form og størrelse af ROI’erne inden for et raster effekten af cirkularitet, fordi grænserne for disse ROI’er ikke påvirkes af støj (Kriegeskorte et al., 2009). Desuden letter denne metode kvantitativ sammenligning af resultater mellem betingelser og potentielt mellem forskellige undersøgelser, hvilket letter den kvantitative reproducerbarhed af fMRI-resultater. Placering af ROI’erne over de lokale maksima optimerede effekt. Det er imidlertid kendt, at aktivitetstoppets placering er påvirket af støj, og derfor kan den nøjagtige placering af ROI’erne altid reproduceres for hver enkelt ROI. Dette påvirker ikke netværksresultaterne, da alle ROI’er er gennemsnit, og den nøjagtige placering er derfor ikke så relevant.
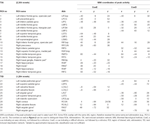
Tabel 1. Aktivitetsmønster.
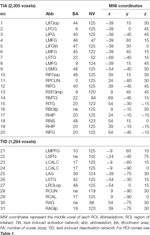
Tabel 2. Region of interest (ROI) karakteristika.
ROI’er over regioner med en stigning i signalændring blev kombineret i et opgaveinduceret aktiveringsnetværk (“TIA”). ROI’er over regioner med et fald i signalændring blev kombineret i et opgaveinduceret deaktiveringsnetværk (“TID”; Tabellerne 1, 2). Hvor det var muligt, blev ROI-navnene bestemt ved hjælp af AAL-atlaset (Tzourio-Mazoyer et al., 2002).
Tabel 2 og figur 2 giver en oversigt over størrelsen og placeringen af alle ROI’er. Fordi størrelsen og grænserne for ROI’erne er foruddefineret, er det muligt, at den gennemsnitlige aktivitet i ROI’en ikke er signifikant forskellig fra baseline, f.eks. fordi antallet af aktiverede voxels inden for ROI’en er relativt lille. Selv om den gennemsnitlige aktivitet på netværksniveau er signifikant forskellig fra baseline for alle betingelser (tabel 3), er dette ikke tilfældet for otte af de 31 individuelle ROI’er for CueE og tre af 31 ROI’er for CueD (tabel 4). Kun voxels, der indeholdt signal for hver deltager, blev inkluderet i analysen. Den maksimale størrelse af en ROI er 125 voxels, nogle ROI’er omfatter mindre end 125 voxels, fordi de er placeret tæt på kraniet.
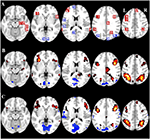
Figur 2. Oversigt over placering af region af interesse (ROI) og fMRI-resultater for hele hjernen. (A) ROI’er for opgaveinduceret aktivering (rød) og opgaveinduceret deaktivering (blå) Bemærk: ROI 22 vises ikke, fordi placeringen er uden for de valgte skiver. MNI-koordinater, BA-nummer og størrelse af ROI’erne kan findes i tabel 2. (B) Visuel repræsentation af t-værdierne for CueE vs. baseline (≥3 = rød til gul, ≤ -3 = blå til lyseblå), (C) T-værdierne for CueD vs. baseline (≥3 = rød til gul, ≤ -3 = blå til lyseblå). Billederne er i neurologisk orientering (L = venstre, R = højre), navne, forkortelser, MNI-koordinater for peak-aktiveringsvoxelerne findes i tabel 1.

Tabel 3. Netværksresultater.
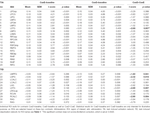
Tabel 4. ROI-resultater.
Test af hypoteser
Vi testede følgende hypoteser i denne undersøgelse:
Hypotese 1: TIA vil vise øget signalændring for CueD, sammenlignet med CueE.
Hypotese 2: TID vil vise nedsat signalændring for CueD, sammenlignet med CueE.
Statistisk analyse
Magtanalyse blev ikke udført, da det var første gang, at dette specifikke opgavedesign blev præsenteret for deltagerne. Derfor er der ingen viden om den forventede procentvise signalændring og variabilitet af betingelserne (Desmond og Glover, 2002).
Tasknøjagtighed blev beregnet for StimE og StimD som den procentvise andel af korrekte svar. Reaktionstider blev beregnet for StimE og StimD over alle korrekte svar.
Der blev anvendt t-tests med én prøve til at sammenligne den procentvise signalændring for TIA og TID sammenlignet med baseline for CueE og CueD. Parrede ensidige t-tests blev brugt til at sammenligne signalændring, nøjagtighed og reaktionstidsforskelle mellem CueD og CueE (hypotese 1 og 2).
Post hoc-testning involverede separate t-tests for hver ROI inden for hvert netværk. ROI-analysen blev Bonferroni-korrigeret, hvis netværksomnibustesten ikke var signifikant. Vi præsenterer ukorrigerede p-værdier og en kommentar, der angiver, om denne p-værdi ville have overlevet Bonferroni-korrektion for alle multiple sammenligninger. SPSS 24 blev anvendt til alle statistiske analyser.
Resultater
Performance
Nøjagtigheden for de lette stimuli (“StimE”) var 98 % ± 1,2 % SEM og for vanskelige stimuli (“StimD”) 85 % ± 2,6 %. Reaktionstiden for StimE var 682 ± 20 ms og for StimD 1.138 ± 27 ms. Begge betingelser viser tilstrækkelige nøjagtighedsrater til, at man kan være sikker på, at opgaven blev udført som instrueret af alle deltagerne. Som antaget reagerede deltagerne mindre præcist (T = -4,3, p < 0,001) og langsommere (T = 17,8, p < 0,001) for StimD end for StimE (figur 3), hvilket indikerer, at kontrasten mellem de lette og vanskelige stimuli var vellykket.
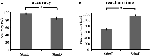
Figur 3. Præstationsresultater. (A) Nøjagtighed for begge stimuluskategorier, (B) reaktionstid for begge stimuluskategorier. Fejlstænger repræsenterer standardfejl i forhold til gennemsnittet (SEM). *p ≤ 0.001. Forkortelser: StimE, et stikord, der angiver en let stimulus, efterfulgt af en let stimulus; StimD, et stikord, der angiver en vanskelig stimulus, efterfulgt af en vanskelig stimulus.
Funktionel MRI
Beskrivelser
TIA-netværket bestod af 20 ROI’er, hvoraf 10 var baseret på lokale maksima for CueE, fem var baseret på lokale maksima for CueD, og fem indeholdt både et CueE- og et CueD-maksimum (tabel 1). Af disse 20 ROI’er viste 19 signifikant aktivitet sammenlignet med baseline for CueE, 18 for CueD (tabel 1). Den gennemsnitlige signalændring i TIA sammenlignet med baseline for CueE var 0.12 ± 0.02 (t = 6.11, p < 0.001) og for CueD 0.12 ± 0.02 (t = 7.61, p < 0.001).
TID-netværket bestod af 11 ROI’er, hvoraf tre var baseret på lokale maksima for CueE, seks var baseret på lokale maksima for CueD, og to indeholdt både et CueE- og et CueD-maksimum (tabel 1). Af disse 11 ROI’er viste fire signifikant aktivitet sammenlignet med baseline for CueE, 10 for CueD (tabel 1). Den gennemsnitlige signalændring i TID sammenlignet med baseline for CueE var -0.07 ± 0.02 (t = 2.68, p < 0.01), og for CueD -0.12 ± 0.02 (t = 5.37, p < 0.001; Tabel 3).
Tuning til den forventede sværhedsgrad
Signalforøgelsen i TIA var ikke signifikant stærkere for CueD sammenlignet med CueE (t = -0.24, p = 0.41), hvilket indikerer, at niveauet af TIA-aktivering ikke er afstemt til den forventede sværhedsgrad (Figur 4, Tabel 3). Post hoc ROI-analyse viste ingen af de 20 ROI’er inden for TIA med en signifikant signalforøgelse for CueD sammenlignet med CueE efter anvendelse af Bonferroni-korrektion (Tabel 4, Figur 5).
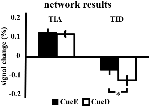
Figur 4. Oversigt over netværksresultaterne. Fejlstænger repræsenterer SEM. *p ≤ 0,05 for cue vanskeligt vs. cue let. Forkortelser: TIA, opgaveinduceret aktiveringsnetværk; TID, opgaveinduceret deaktiveringsnetværk; CueE, et cue, der angiver en let stimulus, ikke efterfulgt af en stimulus; CueD, et cue, der angiver en vanskelig stimulus, ikke efterfulgt af en stimulus.
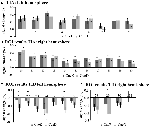
Figur 5. Oversigt over individuelle ROI-svar inden for det opgaveinducerede aktiveringsnetværk, venstre (A) og højre hemisfære (B), og opgaveinduceret deaktivering, venstre (C) og højre hemisfære (D). Fejlstænger repræsenterer SEM. *p ≤ 0,05 for CueD vs. CueE, se figur 2 for placering af ROI’erne, se tabel 1 for navne, forkortelser og MNI-koordinater for ROI’erne.
Signalfaldet i TID var signifikant stærkere for CueD sammenlignet med CueE (T = -2,48, p = 0,01), hvilket indikerer, at niveauet af TID er afstemt efter den forventede sværhedsgrad (tabel 3, figur 4). Post hoc ROI-analyse viste fire ud af 11 ROI’er af TID med et signifikant signalfald for CueD sammenlignet med CueE, nemlig LMPFG (T = -1.83, p = 0.04), LSPG (T = -2.32, p = 0.02), LSTG (T = -2.69, p < 0.01), og RCALC (T = -2.90, p < 0.01). Disse resultater indikerer, at tuning til den forventede vanskelighed er stærkest i venstre frontale, temporale og visuelle ROI’er (tabel 4, figur 5).
Diskussion
I denne undersøgelse undersøgte vi, om og hvordan fasisk årvågenhed er tunet til den forventede vanskelighed af en kommende kognitiv udfordring. Phasisk årvågenhed blev afspejlet i aktiverede såvel som deaktiverede hjerneområder. Det aktiverede netværk omfattede regioner i det centrale udøvende netværk. Det deaktiverede netværk omfattede regioner i DMN samt visuelle cortexer. Niveauet af deaktivering var proportionalt med den forventede sværhedsgrad af den kommende stimulus, mens niveauet af aktivering ikke var det. Disse resultater tyder på, at modulation af fasisk årvågenhed overvejende afspejles i graduerede undertrykkelsesprocesser, der er irrelevante for den aktuelle opgave (f.eks. kardio-vagale, auditive og visuelle processer). Muligvis skete dette, fordi disse processer kunne forstyrre kommende kognitive udfordringer.
Forrige billeddannelsesundersøgelser af fasisk årvågenhed brugte ét sværhedsniveau, præsenterede kun aktiverede hjerneområder (Shulman et al., 1999; Sturm og Willmes, 2001; Thiel et al., 2004; Fan et al., 2005; Périn et al., 2010; Yanaka et al., 2010), eller fokuserede udelukkende på aktivitet i visuel cortex (Bartolucci og Smith, 2011). Disse undersøgelser har identificeret et overvejende højre-lateraliseret fronto-parietal-thalamisk netværk, undertiden kombineret med motoriske og visuelle kortikale regioner, der blev aktiveret af et cue. Der er væsentlige forskelle i de aktiverede netværk mellem undersøgelserne. Nogle af forskellene mellem netværkene i tidligere undersøgelser kan forklares med den opgave, der fulgte efter cue’et. For eksempel identificerede Thiel et al. (2004) kun aktivering i visuelle kortikale regioner ved hjælp af en visuel opfattelsesopgave, mens Fan et al. (2005) og Yanaka et al. (2010) identificerede thalamisk og motorisk aktivering ved hjælp af en motorisk responsopgave. Selv om Bartolucci og Smith (2011) præsenterer et elegant cued opgavedesign med fire sværhedsgrader af en orienteringsdiskriminationsopgave, præsenterede de kun aktivering i den visuelle cortex. Derfor er det uklart, hvordan andre hjerneområder reagerede i denne opgave. Det opgaveinducerede aktiveringsnetværk, der blev identificeret i den aktuelle undersøgelse, ligner mest det netværk, der blev fundet af Shulman et al. (1999), der brugte en lignende type visuel behandlingsopgave.
Det er uklart, om fasisk årvågenhed er et bottom-up eller top-down respons på et uinformativt cue (Thiel et al., 2004; Hackley, 2009; Périn et al., 2010; Bartolucci og Smith, 2011; Chica et al., 2016). Fasisk årvågenhed er af nogle blevet betragtet som en bottom-up (Sturm et al., 1999; Hackley, 2009), af andre som top-down (Bartolucci og Smith, 2011), og af andre som en kombination (Thiel et al., 2004; Périn et al., 2010) respons. Vi argumenterer for, at det er mest hensigtsmæssigt at betegne hjernens respons på et cue som en kombination af bottom-up og top-down behandling, fordi deltagerne både reagerer på et eksternt cue og en instruktion forbundet med dette cue. Vores vigtigste resultater vedrører imidlertid forskellen i respons på to lignende cues, der angiver en anden type opgavestimulus. Derfor argumenterer vi for, at den resulterende forskel mellem det cue, der indikerer en vanskelig opgave, og det cue, der indikerer en let opgave, kan repræsentere et niveau af top-down kontrol relateret til fortolkningen af cue og den tilknyttede instruktion.
Det opgaveinducerede deaktiveringsnetværk omfattede hjerneområder i DMN, nemlig den mediale præfrontale gyrus, superior temporal gyri og angulære gyri (Shulman et al., 1999; Raichle et al., 2001; Raichle, 2015). Det er almindeligvis en hypotese, at DMN-deaktivering hjælper opgaveudførelsen ved at reducere interne processer, der forstyrrer kognitive udfordringer (Raichle et al., 2001; Buckner et al., 2008; Raichle, 2015). Dette synspunkt understøttes af undersøgelser, der viser, at niveauet af DMN-deaktivering skalerer med opgavens sværhedsgrad under udførelsen af opgaven (McKiernan et al., 2003; Singh og Fawcett, 2008). Disse undersøgelser undersøgte deaktivering under opgaveudførelse. Vores resultater tyder på, at denne skalering allerede kan forekomme under opgaveforventningsfasen. Weissman et al. (2006) undersøgte variabiliteten i de individuelle reaktionstider for at identificere lapses of attention. De manglende opmærksomhed kunne være forbundet med reduceret DMN-deaktivering. Mens (Weissman et al., 2006) kiggede på naturlig variation i opmærksomhedsniveauet, viser vores resultater, at deaktivering i flere DMN-regioner også kan indstilles på en top-down måde, før opgavestimulus præsenteres.
De mest dybtgående indstillingsvirkninger blev fundet i de venstre mediale præfrontale, temporale og visuelle regioner. Nedenfor diskuterer vi mulige implikationer af disse effekter for disse regioner i forhold til deres foreslåede funktion.
Den mediale præfrontale cortex-aktivitet er tidligere blevet forbundet med mind-wandering (Bertossi et al., 2017), hukommelseshentning (Euston et al., 2012) og kardiovagal kontrol (Wong et al., 2007). Denne mediale præfrontale deaktivering tyder på, at en af disse funktioner allerede er undertrykt i forventning om en kognitiv udfordring.
Den tuning af deaktivering til den forventede vanskelighed i visuelle og temporale regioner kan være relateret til undertrykkelse af irrelevant visuelt og auditivt sensorisk input. Afstemningen i de bilaterale temporale regioner kan specifikt være relateret til undertrykkelse af den støj, der genereres af MRI-skanneren. Det er rimeligt at forvente, at lydniveauet er ens mellem de forskellige betingelser. Det forhold, at deaktiveringsniveauet er stærkere for de vanskelige cues, tyder på, at behandlingen af omgivende lyd undertrykkes stærkere, hvis der forventes en vanskelig opgave. Deaktivering i visuelle cortexer er mindre almindeligt rapporteret end deaktivering i de temporale regioner. Smith et al. (2000) har imidlertid tidligere rapporteret en sammenhæng mellem udbredt deaktivering i den visuelle cortex og opmærksomhed på en specifik del af et billede. Giesbrecht et al. (2006) rapporterede også om modulation af visuel aktivering i forbindelse med skjult opmærksomhed på objekter nær midten af deres synsfelt sammenlignet med objekter i det perifere synsfelt. Disse resultater tyder på, at aktiveringen i de visuelle cortexer kan moduleres af skjult opmærksomhed på enten foveale eller perifere regioner. Vores undersøgelse tilføjer en ny dimension til disse fund, da den antyder, at aktivering i de visuelle cortexer ikke kun moduleres af placeringen af skjult visuel opmærksomhed, men også af den forventede sværhedsgrad af en visuel udfordring; og da signalerne var identiske i placering og størrelse synes denne proces at være reguleret på en top-down måde.
Vi identificerede også et sæt hjerneområder, der viste øget aktivering i forventning om en kognitiv udfordring. Dette netværk viste lighed med det centrale udøvende netværk, der tidligere er blevet forbundet med udførelsen af arbejdshukommelsesopgaver (Lawrence et al., 2003). Aktiveringen i dette netværk var imidlertid ikke indstillet på den forventede opgavesværhedsgrad. Dette tyder på, at disse regioner repræsenterer processer, der er ens for den vanskelige og lette betingelse. Muligvis er aktivitet i dette netværk forbundet med evaluering af cue eller genfinding og vedligeholdelse af opgavens kontekst og instruktioner.
Der skal tages hensyn til nogle begrænsninger, der kan påvirke fortolkningen af vores resultater. For det første inkluderede vi ikke forsøg med stimuli, der ikke blev forudgået af et cue, og derfor er det ikke muligt at undersøge effekten af cue’et på kognitiv præstation. På grund af tidsbegrænsning var det ikke muligt at gentage vores opgave med identiske stimuli, men uden cues.
For det andet brugte vi et udråbstegn som en del af cueet, der indikerer en vanskelig kognitiv udfordring, fordi dette symbol typisk bruges, hvis der er behov for øget årvågenhed. Det er muligt, at udråbstegnet i sig selv kan have spillet en rolle i de forskellige aktiveringsmønstre for den lette og den vanskelige tilstand på grund af dets iboende betydning. Dette påvirker dog ikke hovedfortolkningen af vores resultater, bortset fra at vores hovedresultater kan klassificeres som en bottom-up-reaktion i stedet for en top-down-reaktion. Endelig giver vores eksperimentelle design os ikke mulighed for at undersøge, om aktiviteten relateret til fasisk årvågenhed kan generaliseres til andre domæner, eller om den er domænespecifik, da den kun omfattede en visuo-perceptuel kognitiv udfordring.
For at konkludere har vi i denne undersøgelse vist, at afstemningen af fasisk årvågenhed er repræsenteret ved niveauet af deaktivering i flere regioner. Denne effekt er stærkest i den mediale præfrontale, visuelle og temporale cortex, hvilket kan afspejle en undertrykkelse af kardiovagal kontrol, visuel behandling i det perifere synsfelt og undertrykkelse af MRI-scannerstøj. Disse resultater tyder på, at kognitiv præstation fremmes af en tilstand i hjernen, der er tæt koblet til forventninger om sværhedsgraden af en kommende kognitiv udfordring.
Datatilgængelighed
Rå data blev genereret på Elisabeth-Tweesteden-hospitalet. Afledte data, der understøtter resultaterne af denne undersøgelse, er tilgængelige fra den tilsvarende forfatter efter rimelig anmodning.
Ethisk erklæring
Denne undersøgelse blev udført i overensstemmelse med anbefalingerne fra den hollandske lov om social støtte, medicinsk etisk komité Brabant. Alle forsøgspersoner gav skriftligt informeret samtykke i overensstemmelse med Helsinki-erklæringen. Protokollen blev godkendt af den medicinske etiske komité Brabant.
Author Contributions
MD og JJ skrev hovedmanuskriptet og udarbejdede figurerne. MD og IS udførte eksperimenterne. G-JR, NR og JJ førte i fællesskab tilsyn med arbejdet. Alle forfattere gennemgik manuskriptet.
Funding
Denne undersøgelse er blevet finansieret af ZonMW, en hollandsk national organisation (Netherlands Organization for Health Research and Development, ZONMW; projektnummer: 842003004), projekttitel:
Interessekonflikter
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller økonomiske relationer, der kunne opfattes som en potentiel interessekonflikt.
Akkreditering
Vi vil gerne takke radiograferne William Pigmans og Maikel Brands for deres bidrag til denne undersøgelse.
Abkortninger
DMN, default mode network; M, mand; F, kvinde; GLM, generel lineær model; SPM, statistisk parametrisk kortlægning; CueE, et cue, der angiver en let stimulus, ikke efterfulgt af en stimulus; CueD, et cue, der angiver en svær stimulus, ikke efterfulgt af en stimulus; StimE, et cue, der angiver en let stimulus, efterfulgt af en let stimulus; StimD, et cue, der angiver en vanskelig stimulus, efterfulgt af en vanskelig stimulus; MRI, magnetisk resonansbilleddannelse; ROI, region af interesse; GDL, GNU data language; TIA, opgaveinduceret aktiveringsnetværk; TID, opgaveinduceret deaktiveringsnetværk; SEM, standard error of the mean.
Fodnoter
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M., og Smith, A. T. (2011). Opmærksomhedsmodulation i visuel cortex ændres under perceptuel læring. Neuropsychologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi, E., Peccenini, L., Solmi, A., Avenanti, A. og Ciaramelli, E. (2017). Transkraniel jævnstrøms stimulation af den mediale præfrontale cortex dæmper tankeløshed hos mænd. Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., og Schacter, D. L. (2008). Hjernens standardnetværk: anatomi, funktion og relevans for sygdom. Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Buracas, G. T., og Boynton, G. M. (2002). Effektivt design af begivenhedsrelaterede fMRI-eksperimenter ved hjælp af M-sekvenser. Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | CrossRef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P., og Bushnell, M. C. (2015). Er et responsivt standardtilstandsnetværk påkrævet for succesfuld arbejdshukommelsesopgaveudførelse? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., og Paz-Alonso, P. M. (2016). Interaktioner mellem fasisk alarmering og bevidsthed i det fronto-striatale netværk. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | CrossRef Full Text | Google Scholar
Desmond, J. E., og Glover, G. H. (2002). Estimering af stikprøvestørrelsen i funktionelle MRI (fMRI) neuroimaging-undersøgelser: statistiske styrkeanalyser. J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., og McNaughton, B. L. (2012). Den mediale præfrontale cortex’ rolle i hukommelse og beslutningstagning. Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., og Posner, M. I. (2005). Aktivering af opmærksomhedsnetværk. Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., og Posner, M. I. (2002). Test af effektiviteten og uafhængigheden af opmærksomhedsnetværk. J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/08989892902317361886
PubMed Abstract | CrossRef Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G., og Mangun, G. R. (2006). Pre-målaktivitet i visuel cortex forudsiger adfærdsmæssig præstation på rumlige og funktionelle opmærksomhedsopgaver. Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009). Fremskyndelse af frivillig reaktion ved et advarselssignal. Psychophysiology 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R., Sorsa, M., og Siivonen, P. (2008). Virkninger af 6/6- og 4/8-vagtsystemer på søvnighed blandt broofficerer. Chronobiol. Int. 25, 413-423. doi: 10.1080/0742052020802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A., og Buckner, R. L. (2012). Manglende modulering af opmærksomhedskontrol i avanceret aldring forbundet med patologi i hvidt stof. Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., og Duyn, J. H. (2007). fMRI-undersøgelse af indsats og informationsbehandling i en arbejdshukommelsesopgave. Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., og Rutten, G. J. (2017). “A proposal for a system that facilitates quantitative replication of fMRI studies,” in Proceedings of the Human Brain Mapping Conference 2017 June 25-29, Vancouver, Canada. Tilgængelig online på: https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., og Baker, C. I. (2009). Cirkulær analyse i systemneurovidenskab: farerne ved dobbeltdykning. Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A., og Bartolomeo, P. (2011). Phasisk auditiv alarmering forbedrer visuel bevidst opfattelse. Bevidst. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G., og Stein, E. S. (2003). Flere neurale netværk medierer vedvarende opmærksomhed. J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/08989892903770007416
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, G., Sobering, G., Duyn, J., og Moonen, C. T. (1993). En funktionel MRI-teknik, der kombinerer principperne for ekkoforskydning med en række observationer (PRESTO). Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | CrossRef Full Text | Google Scholar
Macleod, J. W., Lawrence, M. A., McConnell, M. M. M., Eskes, G. A., Klein, R. M., og Shore, D. I. (2010). Vurdering af ANT: psykometriske og teoretiske overvejelser om opmærksomhedsnetværkstesten. Neuropsychology 24, 637-651. doi: 10.1037/a0019803
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., D’Angelo, B. R. D. D., Kaufman, J. N., and Binder, J. R. (2006). Afbrydelse af “stream of consciousness”: en fMRI-undersøgelse. Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., Kaufman, J. N., Kaufman, J. N., Kucera-Thompson, J. K. og Binder, J. R. (2003). En parametrisk manipulation af faktorer, der påvirker opgaveinduceret deaktivering i funktionel neuroimaging. J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/08989892903321593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R. D., og Brady, C. B. (1993). Fasisk og tonisk årvågenhed ved Alzheimers sygdom. Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., og Ramsey, N. F. (2008). Forbedret følsomhed med hurtig tredimensionel blod-syresyre-niveau-afhængig funktionel MRI: sammenligning af SENSE-PRESTO og 2D-EPI ved 3 T. NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., og de Marco, G. (2010). Alarmberedskab hos unge raske personer: en fMRI-undersøgelse af hjerneområdeinteraktivitet, der forstærkes af et advarselssignal. Brain Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H., et al. (2009). Virkning af arbejdshukommelsesbelastning på FMRI hvilestatusmønster i efterfølgende hvilefaser. PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E. (2015). Hjernens standardtilstandsnetværk. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E., MacLeod, J. W., Snyder, A. Z., Powers, W. J., Gusnard, D. A., og Shulman, G. L. (2001). En standardmodus for hjernefunktion. Proc. Natl. Acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Petersen, S. E., et al. (1999). Områder, der er involveret i kodning og anvendelse af retningsforventninger til bevægelige objekter. J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K. D., og Fawcett, I. P. (2008). Forbigående og lineært gradueret deaktivering af det menneskelige default-mode-netværk ved en visuel detektionsopgave. Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, A. T., Singh, K. D., og Greenlee, M. W. (2000). Opmærksomhedsundertrykkelse af aktivitet i den menneskelige visuelle cortex. Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., de Simone, A., Krause, B. J., Specht, K., Hessemann, V., Radermacher, I., et al. (1999). Funktionel anatomi af intrinsisk årvågenhed: beviser for et froto-parietal-thalamisk-hjernestam-netværk i højre hjernehalvdel. Neuropsychologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., og Willmes, K. (2001). Om den funktionelle neuroanatomi af intrinsisk og fasisk årvågenhed. Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | CrossRef Full Text | Google Scholar
Thiel, C. M., Zilles, K., og Fink, G. R. (2004). Cerebrale korrelater af alarmering, orientering og reorientering af visuospatial opmærksomhed: en begivenhedsrelateret fMRI-undersøgelse. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Automatiseret anatomisk mærkning af aktiveringer i SPM ved hjælp af en makroskopisk anatomisk parcellering af MNI MRI-hjernen for enkeltpersoner. Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | CrossRef Full Text | Google Scholar
van Gelderen, P., Ramsey, N. F., Liu, G., Duyn, J. H., Frank, J. A., Weinberger, D. R., et al. (1995). Tre-dimensionel funktionel magnetisk resonansafbildning af den menneskelige hjerne på en klinisk 1,5-T scanner. Proc. Natl. Acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N., og Henik, A. (2011). Fasisk årvågenhed kan modulere eksekutiv kontrol ved at forbedre den globale behandling af visuelle stimuli. Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., Visscher, K. M., og Woldorff, M. G. (2006). De neurale baser for momentane lapses i opmærksomhed. Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | CrossRef Full Text | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S., og Shoemaker, J. K. (2007). Ventral medial præfrontal cortex og kardiovagal kontrol hos bevidste mennesker. Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka, H. T., Saito, D. N., Uchiyama, Y., og Sadato, N. (2010). Neurale substrater af fasisk årvågenhed: en funktionel magnetisk resonansbilleddannelsesundersøgelse. Neurosci. Res. 68, 51-58. doi: 10.1016/j.neures.2010.05.005
PubMed Abstract | CrossRef Full Text | Google Scholar