Introducere
Cercetarea anterioară arată că oamenii pot folosi un indiciu pentru a se pregăti mental pentru o provocare cognitivă. Dacă participanții sunt instruiți să se pregătească mental în momentul în care este prezentat un indiciu, acest lucru a fost definit ca alertă fazică . Capacitatea de a ne influența nivelul de vigilență ar putea fi importantă, în special dacă vigilența vine și cu un cost. Dacă solicitarea de vigilență este prea mare, aceasta ar putea duce la oboseală (Härmä et al., 2008) sau la probleme legate de stres. În acest studiu, ne propunem să examinăm dacă și modul în care adulții sănătoși își adaptează nivelul de vigilență la dificultatea preconizată a unei sarcini cognitive viitoare.
Dacă un indiciu este prezentat chiar înainte de un stimul al sarcinii, oamenii sunt, în general, capabili să răspundă mai repede în sarcini simple de răspuns motor (Fan et al., 2002; Macleod et al., 2010; Weinbach și Henik, 2011) și să perceapă stimulii vizuali degradați cu mai multă acuratețe (Kusnir et al., 2011). Studiile imagistice au arătat că vigilența fazică este asociată cu o activare crescută în regiunile cerebrale frontale, parietale, talamice (Shulman et al., 1999; Fan et al., 2005; Yanaka et al., 2010), temporo-occipitale (Thiel et al., 2004) și vizuale (Bartolucci și Smith, 2011). Aceste studii imagistice au furnizat informații valoroase despre reprezentarea vigilenței fazice în creier. Cu toate acestea, vigilența fazică a fost studiată, în general, ca un fenomen on/off sau analiza a fost limitată la cortexul vizual. Prin urmare, nu este clar dacă vigilența reflectă, de asemenea, dificultatea așteptată a provocării cognitive în rețelele cerebrale întregi.
În plus, studiile anterioare au indicat că dezactivarea regiunilor cerebrale care fac parte din rețeaua modului implicit (DMN) poate facilita executarea sarcinilor cognitive. În primul rând, studiile privind executarea sarcinilor arată că un nivel crescând de dificultate a sarcinii este asociat cu o dezactivare tot mai mare (McKiernan et al., 2003, 2006; Jansma et al., 2007; Singh și Fawcett, 2008; Pyka et al., 2009; Hedden et al., 2012; Čeko et al., 2015). În al doilea rând, Jansma et al. (2007) au arătat că partea prefrontală medială a DMN se acordează doar cu dificultatea sarcinii, pe care participantul o putea anticipa, dar nu a fost afectată de dificultatea reală a fiecărui stimul, pe care participantul nu o putea anticipa. În cele din urmă, Weissman et al. (2006) au constatat că dezactivarea în DMN chiar înainte de stimul a fost mai slabă dacă subiecții au răspuns relativ lent. Poate că acești subiecți au răspuns mai încet pentru că au fost temporar mai puțin alerți.
Pentru studiul nostru, am conceput o sarcină de percepție vizuală indicată cu o condiție ușoară și una dificilă. Indicii au furnizat informații despre dificultatea stimulului viitor, dar nu au furnizat informații care să faciliteze sarcina în sine. În jumătate din încercări, indiciul nu a fost urmat de un stimul. Doar aceste încercări au fost analizate pentru activitatea cerebrală, astfel încât am putut izola complet activarea asociată cu indiciul, de activarea asociată cu executarea sarcinii. Am emis ipoteza că participanții își reglează activitatea în funcție de dificultatea așteptată a sarcinii.
Materiale și metode
Participanți
Participanții au fost recrutați prin anunțuri online. Participanții au fost excluși dacă au raportat antecedente de tulburări neurologice sau psihiatrice semnificative, sau contraindicații pentru scanarea prin rezonanță magnetică (RMN) (obiecte metalice în corp sau în jurul acestuia, claustrofobie sau sarcină). Acest studiu a fost realizat în conformitate cu recomandările Legii privind cercetarea medicală care implică subiecți umani (WMO), Comitetul de etică a cercetării medicale Brabant. Protocolul a fost aprobat de Comitetul de etică a cercetării medicale (număr de protocol: NL51147.028.14). Toți subiecții și-au dat consimțământul informat în scris, în conformitate cu Declarația de la Helsinki.
Au participat la studiu 22 de voluntari dreptaci sănătoși. Datele de la doi participanți au fost excluse din cauza artefactelor de scanare. Rezultatele sunt raportate pentru cei 20 de participanți rămași.
Designul sarcinii
Am conceput o sarcină (cu design legat de eveniment) care ne-a permis să examinăm efectele de anticipare legate de dificultatea așteptată a unei sarcini, fără a confunda efectele de execuție a sarcinii.
Stimulii sarcinii au constat din nouă săgeți într-o dispunere de trei pe trei pe un ecran negru (Figura 1). Un „stimul ușor” conținea opt săgeți în direcția corectă și o săgeată în direcția opusă. Un „stimul dificil” conținea cinci săgeți în direcția corectă și patru săgeți în direcția opusă (figura 1). Participanții au fost instruiți să apese un buton cu mâna care corespundea direcției majorității săgeților. Stimulii ușori și dificili au fost prezentați într-o ordine aleatorie. Stimulii de bază au constat într-un ecran negru staționar cu textul: „aveți o pauză de 30 s” în limba olandeză și cu litere albe. Stimulii au fost prezentați cu ajutorul software-ului „prezentare”.
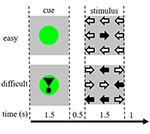
Figura 1. Reprezentarea stimulilor vizuali. Fiecare încercare a fost de 4,5 s. În jumătate din încercări, indiciul nu a fost urmat de un stimul. Aceste încercări au fost utilizate pentru analiza imagisticii prin rezonanță magnetică funcțională (fMRI).
Care încercare a început cu un indiciu care indica dificultatea stimulului prezentat ulterior. Un punct verde a indicat un „stimul ușor”, un punct verde cu semnul exclamării a indicat un „stimul dificil” (Figura 1).
În jumătate din încercări, indiciul nu a fost urmat de un stimul, în cealaltă jumătate stimulul a fost prezentat la 500 ms după ce indiciul a dispărut. Acest lucru a dus la patru condiții: (1) un indiciu pentru un stimul ușor, care nu a fost urmat de un stimul („CueE”); (2) un indiciu pentru un stimul dificil, care nu a fost urmat de un stimul („CueD”); (3) un indiciu pentru un stimul ușor, urmat de un stimul ușor („StimE”); și (4) un indiciu pentru un stimul dificil, urmat de un stimul dificil („StimD”). Toate condițiile au fost comparate cu linia de bază. Rezultatele IRM funcționale (fMRI) s-au bazat pe CueE și CueD. Rezultatele privind performanța s-au bazat pe StimE și StimD. CueE și CueD au fost prezentate într-o ordine pseudo-aleatorie folosind secvențe M pentru a maximiza independența regresorului modelului liniar general (GLM) între condiții și pentru a optimiza eficiența designului (Buracas și Boynton, 2002). Participanții nu au putut prezice dacă un cue va fi urmat sau nu de un stimul.
Durata fiecărui proces a fost de 4.500 ms. Indicii au fost prezentați la t = 0 timp de 1.500 ms, iar stimulii au fost prezentați la t = 2.000, tot timp de 1.500 ms (Figura 1). Sarcina a fost efectuată în două serii de 68 de încercări. Linia de bază a constat în trei perioade de odihnă de 30 s, înainte, după și între cele două runde. Durata totală a sarcinii a fost de 11,7 min. Aceasta a fost a doua sarcină a sesiunii și a început după ce participanții au zăcut aproximativ 10 min în scaner.
Participanții au fost instruiți să se pregătească pentru un stimul ușor sau dificil pe baza informațiilor din indiciu (Figura 1) și să indice cât mai precis și mai rapid posibil direcția majorității săgeților din stimul. Participanții au exersat sarcina în afara scanerului, urmând un protocol de practică standard de 3 min. Acest protocol a fost repetat în cazul în care performanța a fost insuficientă (sub 70% acuratețe pentru stimulii dificili).
O oglindă atașată la bobina capului a permis participanților să vadă un ecran de proiecție transparent poziționat în spatele capului. Un proiector video din interiorul camerei de scanare a proiectat stimulii sarcinii pe ecran. Două cutii de butoane pneumatice cu presiune de aer au fost utilizate pentru a înregistra răspunsurile. Participanții au folosit degetul mare sau arătătorul stâng pentru a răspunde „stânga”, iar cel drept pentru a răspunde „dreapta.”
Achiziționarea imaginilor
Scanările au fost efectuate pe un scaner 3T Philips Achieva (Philips Medical Systems, Best, Țările de Jos) folosind o bobină de cap SENSE cu 32 de canale. O imagine structurală 3D ponderată T1 a fost achiziționată în scopul înregistrării anatomice . imaginile fMRI au fost obținute utilizând o secvență de impulsuri 3D PRESTO . Șase scanări fictive au fost achiziționate și apoi au fost aruncate de către scaner.
Preprocesarea imaginilor
Datele fMRI au fost preprocesate utilizând cartografierea parametrică statistică (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, Londra, UK2). Scanările de la o sesiune au fost realiniate la prima scanare pentru a corecta mișcarea subiectului folosind o abordare prin metoda celor mai mici pătrate, o transformare spațială cu șase parametri (corp rigid) și o estimare B-spline de gradul 2. Scanările au fost coreglate la T1 utilizând un model de corp rigid. Parametrii au fost estimați prin funcția de informație reciprocă normalizată. Imaginile au fost redecupate cu ajutorul unei B-spline de gradul 4. T1 a fost normalizat din punct de vedere spațial în spațiul MNI standard utilizând o regularizare foarte ușoară a polarizării (0,0001) și o B-Spline de gradul 4. Parametrii rezultați au fost aplicați tuturor scanărilor funcționale pentru a minimiza diferențele anatomice și, prin urmare, pentru a permite analiza de grup. În cele din urmă, toate scanările au fost netezite spațial cu un filtru 3D Gaussian (lățimea completă la jumătate maximă: 8 mm) pentru a minimiza și mai mult efectul diferențelor anatomice funcționale.
Analiză fMRI individuală
Analiza de regresie GLM legată de evenimente a fost efectuată pentru fiecare voxel pentru a genera hărți de activare individuală folosind o abordare univariată în masă cu un model de autocorelație globală aproximativă AR(1) și un filtru de trecere înaltă (cut-off de 128 s). Linia de bază nu a fost modelată în mod explicit. Funcția de bază a fost o HRF canonică fără derivate. A fost utilizat un prag de mascare de 0,8. Au fost utilizați regresori separați pentru fiecare condiție (CueE, 17 puncte de timp; CueD, 16 puncte de timp; StimE, 17 puncte de timp; StimD, 18 puncte de timp; și un regresor perturbator pentru perioadele albe în sarcina legată de secvența m, 68 puncte de timp). Niciun punct de timp nu a fost exclus din analiză. Hărțile beta au fost transformate pentru a reflecta schimbarea reală a semnalului procentual în fiecare voxel. Prezentăm doar rezultatele pentru regresorii CueE și CueD, reflectând schimbările de semnal pentru indicii fără stimul. Beta și hărțile t statistice au fost verificate vizual pentru artefacte majore.
Analiză fMRI de grup
A fost efectuată o analiză fMRI de al doilea nivel cu regresie liniară la fiecare voxel pentru vizualizare și selectarea regiunii de interes (ROI). O analiză ROI a fost efectuată cu limbajul de date GNU (GDL3), utilizând hărțile de schimbare a semnalului procentual al subiectului individual generate de analiza GLM. Voxele din cadrul unui ROI sau al unei rețele au fost mediate pentru a calcula diferența de semnal între CueE, CueD și linia de bază.
Selecția ROI
Maximele și minimele locale supratentoriale au fost determinate atât pentru CueE, cât și pentru CueD prin SPM cu un prag necorectat (p < 0,001; Tabelul 1). ROI-uri cubice de dimensiuni și forme predefinite au fost plasate peste maximele locale pentru contrastul CueE-baseline și CueD-baseline separat în cadrul unui raster predefinit (Jansma și Rutten, 2017; Tabelul 2). ROI-urile au fost incluse în analiză dacă au conținut un maxim local semnificativ pentru CueE sau CueD. Prin definirea ROI-urilor pe maximele locale pentru ambele condiții separat, prevenim prejudecata către CueE sau CueD. În plus, forma și dimensiunea predefinite ale ROI-urilor în cadrul unui raster reduc efectul de circularitate, deoarece marginile acestor ROI-uri nu sunt afectate de zgomot (Kriegeskorte et al., 2009). În plus, această metodă facilitează compararea cantitativă a rezultatelor între condiții și, potențial, între diferite studii, facilitând astfel reproductibilitatea cantitativă a rezultatelor fMRI. Plasarea ROI-urilor peste maximele locale a optimizat puterea. Cu toate acestea, se știe că locația vârfului de activitate este afectată de zgomot, prin urmare, locația exactă a ROI-urilor se poate reproduce întotdeauna pentru fiecare ROI. Acest lucru nu afectează rezultatele rețelei, deoarece toate ROI-urile sunt mediate și, prin urmare, locația exactă nu este la fel de relevantă.
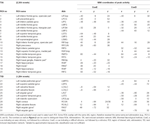
Tabelul 1. Model de activitate.
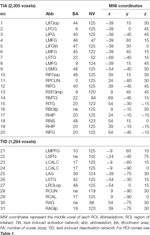
Tabel 2. Caracteristicile regiunii de interes (ROI).
ROI peste regiunile cu o creștere a schimbării semnalului au fost combinate într-o rețea de activare indusă de sarcină („TIA”). ROI-urile peste regiunile cu o scădere a modificării semnalului au fost combinate într-o rețea de dezactivare indusă de sarcină („TID”; tabelele 1, 2). Atunci când a fost posibil, denumirile ROI au fost determinate utilizând atlasul AAL (Tzourio-Mazoyer et al., 2002).
Tabelul 2 și Figura 2 oferă o imagine de ansamblu a dimensiunii și localizării tuturor ROI-urilor. Deoarece dimensiunea și granițele ROI-urilor sunt predefinite, este posibil ca activitatea medie din ROI să nu fie semnificativ diferită față de linia de bază, de exemplu, deoarece numărul de voxeli activați în cadrul ROI-ului este relativ mic. Deși activitatea medie la nivel de rețea este semnificativ diferită față de linia de bază pentru toate condițiile (tabelul 3), acest lucru nu este valabil pentru opt dintre cele 31 de ROI-uri individuale pentru CueE și pentru trei dintre cele 31 de ROI-uri pentru CueD (tabelul 4). Doar voxele care au conținut semnal pentru fiecare participant au fost incluse în analiză. Dimensiunea maximă a unui ROI este de 125 de voxeli, unele ROI-uri includ mai puțin de 125 de voxeli deoarece sunt poziționate în apropierea craniului.
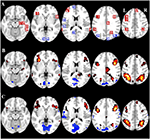
Figura 2. Prezentare generală a localizării regiunii de interes (ROI) și a rezultatelor fMRI la nivelul întregului creier. (A) ROI pentru activarea indusă de sarcină (roșu) și dezactivarea indusă de sarcină (albastru) notă: ROI 22 nu este afișat deoarece locația se află în afara feliilor alese. Coordonatele MNI, numărul BA și dimensiunea ROI-urilor pot fi găsite în Tabelul 2. (B) Reprezentarea vizuală a valorilor t pentru CueE față de linia de bază (≥3 = de la roșu la galben, ≤ -3 = de la albastru la albastru deschis), (C) Valorile T pentru CueD față de linia de bază (≥3 = de la roșu la galben, ≤ -3 = de la albastru la albastru deschis). Imaginile sunt în orientare neurologică (L = stânga, R = dreapta), numele, abrevierile, coordonatele MNI ale voxelului cu activări maxime pot fi găsite în Tabelul 1.

Tabel 3. Rezultatele rețelei.
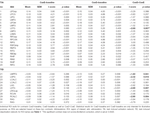
Tabel 4. Rezultatele ROI.
Testarea ipotezelor
Am testat următoarele ipoteze în acest studiu:
Ipoteza 1: TIA va arăta o schimbare de semnal mai mare pentru CueD, în comparație cu CueE.
Ipoteza 2: TID va prezenta o schimbare de semnal diminuată pentru CueD, în comparație cu CueE.
Analiză statistică
Analiza puterii nu a fost efectuată, deoarece a fost prima dată când acest design specific al sarcinii a fost prezentat participanților. Prin urmare, nu se cunoaște procentul așteptat de schimbare a semnalului și variabilitatea condițiilor (Desmond și Glover, 2002).
Precizia sarcinii a fost calculată pentru StimE și StimD ca procent de răspunsuri corecte. Timpii de reacție au fost calculați pentru StimE și StimD pe toate răspunsurile corecte.
S-au folosit teste t cu o singură probă pentru a compara modificarea procentuală a semnalului pentru TIA și TID în comparație cu linia de bază pentru CueE și CueD. S-au folosit teste t cu o singură probă perechi unilaterale pentru a compara schimbarea semnalului, acuratețea și diferențele de timp de reacție între CueD și CueE (Ipoteza 1 și 2).
Testarea post hoc a implicat teste t separate pentru fiecare ROI din cadrul fiecărei rețele. Analiza ROI a fost corectată de Bonferroni dacă testul omnibus al rețelei nu a fost semnificativ. Prezentăm valorile p necorectate și un comentariu care precizează dacă această valoare p ar fi supraviețuit corecției Bonferroni pentru toate comparațiile multiple. SPSS 24 a fost utilizat pentru toate analizele statistice.
Rezultate
Performanță
Precizia pentru stimulii ușori („StimE”) a fost de 98% ± 1,2% SEM, iar pentru stimulii dificili („StimD”) de 85% ± 2,6%. Timpul de reacție pentru StimE a fost de 682 ± 20 ms, iar pentru StimD de 1 138 ± 27 ms. Ambele condiții prezintă rate de acuratețe suficiente pentru a fi siguri că sarcina a fost executată conform instrucțiunilor tuturor participanților. Conform ipotezei, participanții au răspuns mai puțin precis (T = -4,3, p < 0,001) și mai lent (T = 17,8, p < 0,001) pentru StimD decât pentru StimE (Figura 3), ceea ce indică faptul că contrastul dintre stimulii ușori și cei dificili a avut succes.
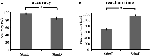
Figura 3. Rezultatele performanței. (A) Precizia pentru ambele categorii de stimuli, (B) timpul de reacție pentru ambele categorii de stimuli. Barele de eroare reprezintă eroarea standard a mediei (SEM). *p ≤ 0.001. Abrevieri: StimE, un indiciu care indică un stimul ușor, urmat de un stimul ușor; StimD, un indiciu care indică un stimul dificil, urmat de un stimul dificil.
RMRI funcțional
Descrieri
Rețeaua TIA a fost alcătuită din 20 de ROI, dintre care 10 s-au bazat pe maxime locale pentru CueE, cinci s-au bazat pe maxime locale pentru CueD, iar cinci conțineau atât un maxim CueE, cât și un maxim CueD (Tabelul 1). Dintre aceste 20 ROI, 19 au prezentat o activitate semnificativă în comparație cu linia de bază pentru CueE, 18 pentru CueD (Tabelul 1). Modificarea medie a semnalului în TIA comparativ cu linia de bază pentru CueE a fost de 0,12 ± 0,02 (t = 6,11, p < 0,001) și pentru CueD de 0,12 ± 0,02 (t = 7,61, p < 0,001).
Rețeaua TID era formată din 11 ROI, dintre care trei se bazau pe maximele locale pentru CueE, șase se bazau pe maximele locale pentru CueD și două conțineau atât un maxim CueE, cât și un maxim CueD (Tabelul 1). Dintre aceste 11 ROI, patru au prezentat o activitate semnificativă în comparație cu linia de bază pentru CueE, 10 pentru CueD (Tabelul 1). Modificarea medie a semnalului în TID comparativ cu linia de bază pentru CueE a fost de -0,07 ± 0,02 (t = 2,68, p < 0,01), iar pentru CueD -0,12 ± 0,02 (t = 5,37, p < 0,001; Tabelul 3).
Tuning la dificultatea așteptată
Creșterea semnalului în TIA nu a fost semnificativ mai puternică pentru CueD în comparație cu CueE (t = -0.24, p = 0.41), ceea ce indică faptul că nivelul de activare a TIA nu este acordat la dificultatea așteptată (Figura 4, Tabelul 3). Analiza ROI post hoc nu a arătat niciunul dintre cele 20 de ROI din cadrul TIA cu o creștere semnificativă a semnalului pentru CueD comparativ cu CueE după aplicarea corecției Bonferroni (Tabelul 4, Figura 5).
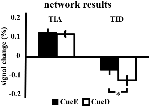
Figura 4. Prezentare generală a rezultatelor rețelei. Barele de eroare reprezintă SEM. *p ≤ 0,05 pentru tacul dificil vs. tacul ușor. Abrevieri: TIA, task induced activation network (rețea de activare indusă de sarcină); TID, task induced deactivation network (rețea de dezactivare indusă de sarcină); CueE, cueE, un cue care indică un stimul ușor, care nu este urmat de un stimul; CueD, un cue care indică un stimul dificil, care nu este urmat de un stimul.
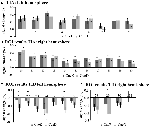
Figura 5. Prezentare generală a răspunsurilor individuale ROI în cadrul rețelei de activare indusă de sarcină, emisfera stângă (A) și emisfera dreaptă (B), și dezactivarea indusă de sarcină, emisfera stângă (C) și emisfera dreaptă (D). Barele de eroare reprezintă SEM. *p ≤ 0.05 pentru CueD vs. CueE, a se vedea Figura 2 pentru localizarea ROI-urilor, a se vedea Tabelul 1 pentru numele, abrevierile și coordonatele MNI ale ROI-urilor.
Diminuarea semnalului în TID a fost semnificativ mai puternică pentru CueD în comparație cu CueE (T = -2.48, p = 0.01), ceea ce indică faptul că nivelul TID este reglat în funcție de dificultatea așteptată (Tabelul 3, Figura 4). Analiza post hoc ROI a arătat patru din 11 ROI ale TID cu o scădere semnificativă a semnalului pentru CueD în comparație cu CueE, și anume LMPFG (T = -1.83, p = 0.04), LSPG (T = -2.32, p = 0.02), LSTG (T = -2.69, p < 0.01) și RCALC (T = -2.90, p < 0.01). Aceste rezultate indică faptul că tuningul la dificultatea așteptată este cel mai puternic în ROI-urile frontale, temporale și vizuale din stânga (Tabelul 4, Figura 5).
Discuție
În acest studiu, am examinat dacă și modul în care vigilența fazică este acordată la dificultatea așteptată a unei provocări cognitive viitoare. Vigilența fazică a fost reflectată în regiunile cerebrale activate, precum și dezactivate. Rețeaua activată a inclus regiuni ale rețelei executive centrale. Rețeaua dezactivată a inclus regiuni ale DMN, precum și cortexuri vizuale. Nivelul de dezactivare a crescut odată cu dificultatea așteptată a stimulului viitor, în timp ce nivelul de activare nu a crescut. Aceste rezultate sugerează că modularea vigilenței fazice se reflectă predominant în procesele de suprimare graduală care sunt irelevante pentru sarcina curentă (de exemplu, procesele cardio-vagale, auditive și vizuale). Este posibil ca acest lucru să se fi întâmplat deoarece aceste procese ar putea interfera cu provocările cognitive viitoare.
Studiile imagistice anterioare privind vigilența fazică au folosit un singur nivel de dificultate, au prezentat doar regiunile cerebrale activate (Shulman et al., 1999; Sturm și Willmes, 2001; Thiel et al., 2004; Fan et al., 2005; Périn et al., 2010; Yanaka et al., 2010) sau s-au concentrat doar pe activitatea din cortexul vizual (Bartolucci și Smith, 2011). Aceste studii au identificat o rețea fronto-parietală-talamică în cea mai mare parte lateralizată spre dreapta, uneori combinată cu regiuni corticale motorii și vizuale, care a fost activată de un indiciu. Există diferențe substanțiale în ceea ce privește rețelele activate între studii. Unele dintre diferențele dintre rețelele din studiile anterioare pot fi explicate prin sarcina care a urmat indicelui. De exemplu, Thiel et al. (2004) au identificat doar activarea în regiunile corticale vizuale folosind o sarcină de percepție vizuală, în timp ce Fan et al. (2005) și Yanaka et al. (2010) au identificat activarea talamică și motorie cu o sarcină de răspuns motor. Deși Bartolucci și Smith (2011) prezintă un design elegant de sarcină cued cu patru niveluri de dificultate a unei sarcini de discriminare a orientării, aceștia au prezentat doar activarea în cortexul vizual. Prin urmare, nu este clar cum au răspuns alte regiuni ale creierului în această sarcină. Rețeaua de activare indusă de sarcină, identificată în studiul actual este cea mai asemănătoare cu rețeaua găsită de Shulman et al. (1999), care a folosit un tip similar de sarcină de procesare vizuală.
Nu este clar dacă vigilența fazică este un răspuns de jos în sus sau de sus în jos la un indiciu neinformativ (Thiel et al., 2004; Hackley, 2009; Périn et al., 2010; Bartolucci și Smith, 2011; Chica et al., 2016). Vigilența fazică a fost considerată de unii ca fiind un răspuns de jos în sus (Sturm et al., 1999; Hackley, 2009), de alții ca fiind de sus în jos (Bartolucci și Smith, 2011), iar de alții ca fiind o combinație (Thiel et al., 2004; Périn et al., 2010). Susținem că este cel mai potrivit să etichetăm răspunsul creierului la un indiciu ca fiind o combinație de procesare de jos în sus și de sus în jos, deoarece participanții răspund atât la un indiciu extern, cât și la o instrucțiune asociată cu acest indiciu. Cu toate acestea, principalele noastre rezultate se referă la diferența de răspuns la două indicii similare care indică un tip diferit de stimul de sarcină. Prin urmare, susținem că diferența rezultată între tacul care indică o sarcină dificilă și tacul care indică o sarcină ușoară poate reprezenta un nivel de control de sus în jos legat de interpretarea tacului și a instrucțiunii asociate.
Rețeaua de dezactivare indusă de sarcină a inclus regiuni cerebrale ale DMN, și anume girusul prefrontal medial, girusul temporal superior și girusul unghiular (Shulman și colab., 1999; Raichle și colab., 2001; Raichle, 2015). Se presupune în mod obișnuit că dezactivarea DMN ajută performanța sarcinilor prin reducerea proceselor interne care interferează cu provocările cognitive (Raichle et al., 2001; Buckner et al., 2008; Raichle, 2015). Această viziune este susținută de studii care arată că nivelul de dezactivare DMN scalează cu dificultatea unei sarcini în timpul executării sarcinii (McKiernan et al., 2003; Singh și Fawcett, 2008). Aceste studii au examinat dezactivarea în timpul executării sarcinii. Rezultatele noastre sugerează că această scalare poate apărea deja în timpul fazei de anticipare a sarcinii. Weissman et al. (2006) au examinat variabilitatea timpilor de reacție individuali pentru a identifica lapsusurile de atenție. Lipsurile de atenție ar putea fi asociate cu o dezactivare redusă a DMN. În timp ce (Weissman et al., 2006) au analizat variația naturală a nivelului de vigilență, rezultatele noastre indică faptul că dezactivarea în mai multe regiuni DMN poate fi, de asemenea, reglată într-o manieră top-down înainte de prezentarea stimulului sarcinii.
Cele mai profunde efecte de reglare au fost găsite în regiunile prefrontală medială, temporală și vizuală din stânga. Mai jos discutăm posibilele implicații ale acestor efecte pentru aceste regiuni în legătură cu funcția lor propusă.
Activitatea cortexului prefrontal medial a fost asociată anterior cu rătăcirea minții (Bertossi și colab., 2017), recuperarea memoriei (Euston și colab., 2012) și controlul cardiovagal (Wong și colab., 2007). Această dezactivare prefrontală medială sugerează că oricare dintre aceste funcții este deja suprimată în anticiparea unei provocări cognitive.
Acordarea dezactivării la dificultatea așteptată în regiunile vizuale și temporale poate fi legată de suprimarea intrărilor senzoriale vizuale și auditive irelevante. Acordarea în regiunile temporale bilaterale poate fi legată în mod specific de suprimarea zgomotului generat de scanerul RMN. Este rezonabil să ne așteptăm ca nivelul sunetului să fie similar între cele două condiții. Faptul că nivelul de dezactivare este mai puternic pentru indicii dificile sugerează că procesarea sunetului ambiental este suprimată mai puternic dacă se așteaptă o sarcină dificilă. Dezactivarea în cortexurile vizuale este mai puțin frecvent raportată decât dezactivarea în regiunile temporale. Cu toate acestea, Smith et al. (2000) au raportat anterior o asociere între dezactivarea generalizată în cortexul vizual și atenția la o anumită parte a unei imagini. Giesbrecht et al. (2006) au raportat, de asemenea, o modulare a activării vizuale asociată cu atenția ascunsă la obiectele din apropierea centrului câmpului lor vizual în comparație cu obiectele din câmpul vizual periferic. Aceste constatări indică faptul că activarea în cortexurile vizuale poate fi modulată de atenția ascunsă fie la regiunile foveale, fie la cele periferice. Studiul nostru adaugă o nouă dimensiune la aceste constatări, deoarece sugerează că activarea în cortexurile vizuale nu este modulată doar de locația atenției vizuale ascunse, ci și de dificultatea așteptată a unei provocări vizuale; și, din moment ce indiciile au fost identice în ceea ce privește locația și dimensiunea, acest proces pare a fi reglementat într-un mod de sus în jos.
Am identificat, de asemenea, un set de regiuni cerebrale care au prezentat o activare crescută în anticiparea unei provocări cognitive. Această rețea a arătat similaritate cu rețeaua executivă centrală care a fost asociată anterior cu executarea sarcinilor de memorie de lucru (Lawrence și colab., 2003). Cu toate acestea, activarea în această rețea nu a fost ajustată la dificultatea așteptată a sarcinii. Acest lucru sugerează că aceste regiuni reprezintă procese care sunt similare pentru condiția dificilă și ușoară. Posibil ca activitatea din această rețea să fie asociată cu evaluarea indiciului sau cu recuperarea și menținerea contextului sarcinii și a instrucțiunilor.
Trebuie luate în considerare unele limitări care pot afecta interpretarea rezultatelor noastre. În primul rând, nu am inclus încercări cu stimuli care nu au fost precedați de un indiciu, prin urmare, nu este posibil să examinăm efectul indicelui asupra performanței cognitive. Din cauza restricției de timp, nu a fost posibil să repetăm sarcina noastră folosind stimuli identici, dar fără indicii.
În al doilea rând, am folosit un semn de exclamare ca parte a indiciului care indică o provocare cognitivă dificilă, deoarece acest simbol este utilizat de obicei dacă este nevoie de o vigilență sporită. Este posibil ca semnul exclamării în sine să fi jucat un rol în diferitele modele de activare pentru condiția ușoară și dificilă, datorită semnificației sale inerente. Cu toate acestea, acest lucru nu afectează interpretarea principală a rezultatelor noastre, cu excepția faptului că rezultatele noastre principale pot fi clasificate ca un răspuns de jos în sus în loc de un răspuns de sus în jos. În cele din urmă, designul nostru experimental nu ne permite să examinăm dacă activitatea legată de vigilența fazică poate fi generalizată la alte domenii sau dacă este specifică domeniului, deoarece a inclus doar o provocare cognitivă visuo-perceptuală.
Pentru a concluziona, în acest studiu am arătat că reglarea vigilenței fazice este reprezentată de nivelul de dezactivare în mai multe regiuni. Acest efect este cel mai puternic în cortexul prefrontal medial, vizual și temporal, ceea ce poate reflecta o suprimare a controlului cardiovagal, procesarea vizuală în câmpul vizual periferic și suprimarea zgomotului scanerului RMN. Aceste rezultate sugerează că performanța cognitivă este facilitată de o stare a creierului care este strâns legată de așteptările privind dificultatea unei provocări cognitive viitoare.
Disponibilitatea datelor
Datele brute au fost generate la spitalul Elisabeth-Tweesteden. Datele derivate care susțin constatările acestui studiu sunt disponibile de la autorul corespondent la o cerere rezonabilă.
Declarație etică
Acest studiu a fost realizat în conformitate cu recomandările legii olandeze de sprijin social, comitetul de etică medicală Brabant. Toți subiecții și-au dat consimțământul informat în scris, în conformitate cu Declarația de la Helsinki. Protocolul a fost aprobat de comitetul de etică medicală Brabant.
Contribuții ale autorilor
MD și JJ au scris manuscrisul principal și au pregătit figurile. MD și IS au efectuat experimentele. G-JR, NR și JJ au supravegheat în comun activitatea. Toți autorii au revizuit manuscrisul.
Finanțare
Acest studiu a fost finanțat de ZonMW, o organizație națională olandeză (Organizația Olandeză pentru Cercetare și Dezvoltare în Sănătate, ZONMW; Număr de proiect: 842003004), titlul proiectului: Implementarea clinică a tehnicilor avansate de RMN pentru localizarea și monitorizarea funcțiilor senzorio-motorii și cognitive la pacienții cu tumori cerebrale.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Am dori să mulțumim radiografilor William Pigmans și Maikel Brands pentru contribuția lor la acest studiu.
Abbreviații
DMN, rețeaua modului implicit; M, bărbat; F, femeie; GLM, model liniar general; SPM, cartografiere parametrică statistică; CueE, un indiciu care indică un stimul ușor, care nu este urmat de un stimul; CueD, un indiciu care indică un stimul dificil, care nu este urmat de un stimul; StimE, un indiciu care indică un stimul ușor, urmat de un stimul ușor; StimD, un indiciu care indică un stimul dificil, urmat de un stimul dificil; RMN, imagistică prin rezonanță magnetică; ROI, regiune de interes; GDL, limbaj de date GNU; TIA, rețea de activare indusă de sarcină; TID, rețea de dezactivare indusă de sarcină; SEM, eroare standard a mediei.
Footnotes
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M., și Smith, A. T. (2011). Modularea atențională în cortexul vizual este modificată în timpul învățării perceptuale. Neuropsychologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi, E., Peccenini, L., Solmi, A., Avenanti, A., și Ciaramelli, E. (2017). Stimularea transcraniană a curentului direct al cortexului prefrontal medial amortizează rătăcirea minții la bărbați. Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | Cross Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R. R., și Schacter, D. L. (2008). Rețeaua implicită a creierului: anatomie, funcție și relevanță pentru boală. Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | Text integral | Google Scholar
Buracas, G. T., și Boynton, G. M. (2002). Proiectarea eficientă a experimentelor fMRI legate de evenimente folosind secvențe M. Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | Refef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P., și Bushnell, M. C. (2015). Este necesară o rețea de mod implicit receptiv pentru o performanță reușită a sarcinilor de memorie de lucru? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., și Paz-Alonso, P. M. (2016). Interacțiuni între alertarea fazică și conștiința în rețeaua fronto-striatal. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | Full Text | Google Scholar
Desmond, J. E., și Glover, G. H. (2002). Estimarea dimensiunii eșantionului în studiile de neuroimagistică IRM funcțională (fMRI): analize de putere statistică. J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | Refef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., și McNaughton, B. L. (2012). Rolul cortexului prefrontal medial în memorie și luarea deciziilor. Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., și Posner, M. I. (2005). Activarea rețelelor atenționale. Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., și Posner, M. I. (2002). Testarea eficienței și independenței rețelelor atenționale. J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/08989290902317361886
PubMed Abstract | Ref Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G., și Mangun, G. R. (2006). Activitatea pre-țintă în cortexul vizual prezice performanța comportamentală pe sarcini de atenție spațială și caracteristică. Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009). Accelerarea reacției voluntare de către un semnal de avertizare. Psychophysiology 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R., Sorsa, M., and Siivonen, P. (2008). Efectele sistemelor de veghe 6/6 și 4/8 asupra somnolenței în rândul ofițerilor de punte. Chronobiol. Int. 25, 413-423. doi: 10.1080/07420520802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A., și Buckner, R. L. (2012). Eșecul de a modula controlul atențional în îmbătrânirea avansată legată de patologia materiei albe. Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | Refef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P. și Duyn, J. H. (2007). studiu fMRI al efortului și al procesării informațiilor într-o sarcină de memorie de lucru. Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | Full Text | Google Scholar
Jansma, J. M., și Rutten, G. J. (2017). „O propunere pentru un sistem care facilitează replicarea cantitativă a studiilor fMRI”, în Proceedings of the Human Brain Mapping Conference 2017, 25-29 iunie, Vancouver, Canada. Disponibil online la adresa: https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., și Baker, C. I. (2009). Analiza circulară în neuroștiința sistemelor: pericolele dublei scufundări. Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | Textul integral | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A. A., și Bartolomeo, P. (2011). Alertarea auditivă fazică auditivă îmbunătățește percepția vizuală conștientă. Conștient. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | Cross Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G., și Stein, E. S. (2003). Rețele neuronale multiple mediază atenția susținută. J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/089892903770007416
PubMed Abstract | Full CrossRef Text | Google Scholar
Liu, G., Sobering, G., Duyn, J. și Moonen, C. T. (1993). O tehnică IRM funcțională care combină principiile de deplasare a ecourilor cu un tren de observații (PRESTO). Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | Text integral | Google Scholar
Macleod, J. W., Lawrence, M. A., McConnell, M. M. M., Eskes, G. A., Klein, R. M., și Shore, D. I. (2010). Aprecierea ANT: considerații psihometrice și teoretice ale testului rețelei de atenție. Neuropsychology 24, 637-651. doi: 10.1037/a001980803
PubMed Abstract | Full CrossRef Full Text | Google Scholar
McKiernan, K. A., D’Angelo, B. R. D., Kaufman, J. N., și Binder, J. R. (2006). Întreruperea „fluxului de conștiință”: o investigație fMRI. Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J. K. K., și Binder, J. R. (2003). O manipulare parametrică a factorilor care afectează dezactivarea indusă de sarcină în neuroimagistica funcțională. J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/089892903321593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R. D., and Brady, C. B. (1993). Vigilența fazică și tonică în boala Alzheimer. Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., și Ramsey, N. F. (2008). Sensibilitate îmbunătățită cu RMN funcțional tridimensional rapid dependent de nivelul de oxigen din sânge: comparație între SENSE-PRESTO și 2D-EPI la 3 T. NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., și de Marco, G. (2010). Starea de alertă la subiecții tineri sănătoși: un studiu fMRI al interactivității regiunilor cerebrale îmbunătățite de un semnal de avertizare. Brain Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H., et al. (2009). Impactul încărcării memoriei de lucru asupra modelului de stare de repaus FMRI în fazele de repaus ulterioare. PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract | Refef Full Text | Google Scholar
Raichle, M. E. (2015). Rețeaua de mod implicit a creierului. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | Red Full Text | Google Scholar
Raichle, M. E., MacLeod, J. W., Snyder, A. Z., Powers, W. J., Gusnard, D. A., și Shulman, G. L. (2001). Un mod implicit de funcționare a creierului. Proc. Natl. acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Petersen, S. E., et al. (1999). Zone implicate în codificarea și aplicarea așteptărilor direcționale la obiectele în mișcare. J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K. D., și Fawcett, I. P. (2008). Dezactivarea tranzitorie și gradată liniar a rețelei umane de mod implicit a modului implicit de către o sarcină de detectare vizuală. Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, A. T., Singh, K. D., și Greenlee, M. W. (2000). Suprimarea atențională a activității în cortexul vizual uman. Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., de Simone, A., Krause, B. J., Specht, K., Hessemann, V., Radermacher, I., et al. (1999). Anatomia funcțională a vigilenței intrinseci: dovezi pentru o rețea froto-parietală-talamică-brainstemică în emisfera dreaptă. Neuropsychologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., și Willmes, K. (2001). Cu privire la neuroanatomia funcțională a vigilenței intrinseci și fazice. Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | Ref Full Text | Google Scholar
Thiel, C. M., Zilles, K., și Fink, G. R. (2004). Corelații cerebrale ale alertei, orientării și reorientării atenției visuospațiale: un studiu fMRI legat de eveniment. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Etichetarea anatomică automatizată a activărilor în SPM folosind o parcelare anatomică macroscopică a creierului MNI MRI cu un singur subiect. Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | Ref Full Text | Google Scholar
van Gelderen, P., Ramsey, N. F., Liu, G., Duyn, J. H., Frank, J. A., Weinberger, D. R., et al. (1995). Imagistica prin rezonanță magnetică funcțională tridimensională a creierului uman pe un scaner clinic de 1,5-T. Proc. Natl. acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N., and Henik, A. (2011). Vigilența fazică poate modula controlul executiv prin îmbunătățirea procesării globale a stimulilor vizuali. Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., K. C., Visscher, K. M., și Woldorff, M. G. (2006). Bazele neuronale ale lapsusurilor momentane în atenție. Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | Textul integral | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S., și Shoemaker, J. K. (2007). Cortexul prefrontal medial ventral și controlul cardiovagal la oamenii conștienți. Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka, H. T., Saito, D. N., Uchiyama, Y., și Sadato, N. (2010). Substratele neuronale ale vigilenței fazice: un studiu de imagistică prin rezonanță magnetică funcțională. Neurosci. Res. 68, 51-58. doi: 10.1016/j.neures.2010.05.005
PubMed Abstract | CrossRef Full Text | Google Scholar
.