Introduction
A korábbi kutatások azt mutatják, hogy az emberek képesek egy jelzést használni arra, hogy mentálisan felkészüljenek egy kognitív kihívásra. Ha a résztvevőket arra utasítják, hogy mentálisan készüljenek fel egy jelző bemutatásának pillanatában, ezt fázisos éberségként definiálták . Az éberségi szintünk befolyásolásának képessége fontos lehet, különösen akkor, ha az éberségnek ára is van. Ha az éberséggel szemben támasztott követelmények túl magasak, az potenciálisan fáradtsághoz (Härmä et al., 2008) vagy stresszel kapcsolatos problémákhoz vezethet. Ebben a tanulmányban azt szeretnénk megvizsgálni, hogy egészséges felnőttek éberségi szintjüket egy közelgő kognitív feladat várható nehézségéhez hangolják-e és hogyan.
Ha egy jelzőt közvetlenül a feladat ingere előtt mutatnak be, az emberek általában gyorsabban képesek reagálni az egyszerű motoros válaszadási feladatokban (Fan et al., 2002; Macleod et al., 2010; Weinbach és Henik, 2011) és pontosabban érzékelik a degradált vizuális ingereket (Kusnir et al., 2011). Képalkotó vizsgálatok kimutatták, hogy a fázisos éberség fokozott aktivációval jár a frontális, parietális, talamikus (Shulman et al., 1999; Fan et al., 2005; Yanaka et al., 2010), temporo-okcipitális (Thiel et al., 2004) és vizuális agyi régiókban (Bartolucci és Smith, 2011). Ezek a képalkotó vizsgálatok értékes információkkal szolgáltak a fázisos éberség agyi reprezentációjáról. A fázisos éberséget azonban általában on/off jelenségként vizsgálták, vagy az elemzést a látókéregre korlátozták. Ezért nem világos, hogy az éberség a kognitív kihívás várható nehézségét is tükrözi-e a teljes agyi hálózatokban.
A korábbi vizsgálatok továbbá azt jelezték, hogy az alapértelmezett üzemmód hálózat (DMN) részét képező agyi régiók deaktiválása megkönnyítheti a kognitív feladatok végrehajtását. Először is, a feladatvégzéssel kapcsolatos vizsgálatok azt mutatják, hogy a feladat növekvő nehézségi szintje növekvő deaktivációval jár együtt (McKiernan et al., 2003, 2006; Jansma et al., 2007; Singh és Fawcett, 2008; Pyka et al., 2009; Hedden et al., 2012; Čeko et al., 2015). Másodszor, Jansma és munkatársai (2007) kimutatták, hogy a DMN mediális prefrontális része csak a feladat nehézségére hangolódik, amelyet a résztvevő előre láthatott, de nem volt hatással az egyes ingerek tényleges nehézsége, amelyet a résztvevő nem láthatott előre. Végül Weissman és munkatársai (2006) azt találták, hogy a DMN-ben közvetlenül az inger előtt a deaktiváció gyengébb volt, ha a kísérleti személyek viszonylag lassan reagáltak. Talán ezek az alanyok azért reagáltak lassabban, mert átmenetileg kevésbé voltak éberek.
A mi vizsgálatunkhoz egy cued vizuális észlelési feladatot terveztünk egy könnyű és egy nehéz feltétellel. A jelzések információt szolgáltattak a közelgő inger nehézségéről, de nem adtak olyan információt, amely magát a feladatot megkönnyítette volna. A próbák felében a jelzést nem követte inger. Csak ezeket a próbákat elemeztük az agyi aktivitás szempontjából, így teljesen el tudtuk különíteni a jelzővel kapcsolatos aktivációt a feladat végrehajtásával kapcsolatos aktivációtól. Feltételezésünk szerint a résztvevők aktivitásukat a feladat várható nehézségéhez hangolják.
Anyagok és módszerek
Tagadók
A résztvevőket online hirdetés útján toboroztuk. A résztvevőket kizárták, ha jelentős neurológiai vagy pszichiátriai rendellenességekről vagy a mágneses rezonancia-vizsgálat (MRI) ellenjavallatairól (fémtárgyak a testben vagy a test körül, klausztrofóbia vagy terhesség) számoltak be. Ezt a vizsgálatot az emberi alanyokat érintő orvosi kutatásokról szóló törvény (WMO), Brabant orvosi kutatási etikai bizottsága ajánlásainak megfelelően végezték. A protokollt az Orvosi Kutatási Etikai Bizottság jóváhagyta (protokollszám: NL51147.028.14). Minden alany a Helsinki Nyilatkozatnak megfelelően írásos beleegyezését adta.
Huszonkét egészséges jobbkezes önkéntes vett részt a vizsgálatban. Két résztvevő adatait kizártuk a szkenner műtárgyak miatt. A fennmaradó 20 résztvevő eredményeit közöljük .
A feladat felépítése
Egy olyan feladatot terveztünk (eseményfüggő tervezéssel), amely lehetővé tette a feladat várható nehézségével kapcsolatos anticipációs hatások vizsgálatát, a feladat végrehajtásának zavaró hatásai nélkül.
A feladat ingerek kilenc nyílból álltak háromszor három elrendezésben egy fekete képernyőn (1. ábra). A “könnyű inger” nyolc nyilat tartalmazott a helyes irányba és egy nyilat az ellenkező irányba. A “nehéz inger” öt nyilat tartalmazott a helyes irányba és négy nyilat az ellenkező irányba (1. ábra). A résztvevőket arra utasították, hogy a nyilak többségének irányának megfelelő kézzel nyomjanak meg egy gombot. A könnyű és nehéz ingereket véletlenszerű sorrendben mutatták be. A kiindulási ingerek egy álló fekete képernyőből álltak a szöveggel: “30 másodperces szünete van” holland nyelven és fehér betűkkel. Az ingereket a “prezentációs” szoftver mutatta be.
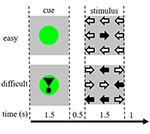
1. ábra. A vizuális ingerek ábrázolása. Az egyes próbák időtartama 4,5 s volt. A próbák felében a jelzést nem követte inger. Ezeket a próbákat használtuk fel a funkcionális mágneses rezonancia képalkotó (fMRI) elemzéshez.
Minden próba a később bemutatott inger nehézségét jelző jelzővel kezdődött. A zöld pont “könnyű ingerre”, a felkiáltójellel ellátott zöld pont “nehéz ingerre” utalt (1. ábra).
A próbák felében a jelzőt nem követte inger, a másik felében az inger a jelző eltűnése után 500 ms-t követően jelent meg. Ez négy körülményt eredményezett: (1) könnyű ingerre vonatkozó jelzés, amelyet nem követett inger (“CueE”); (2) nehéz ingerre vonatkozó jelzés, amelyet nem követett inger (“CueD”); (3) könnyű ingerre vonatkozó jelzés, amelyet könnyű inger követett (“StimE”); és (4) nehéz ingerre vonatkozó jelzés, amelyet nehéz inger követett (“StimD”). Minden körülményt az alaphelyzettel állítottunk szembe. A funkcionális MRI (fMRI) eredmények a CueE és a CueD alapján készültek. A teljesítményeredmények a StimE és a StimD alapján készültek. A CueE-t és a CueD-t pszeudo-randomizált sorrendben mutattuk be M szekvenciák használatával, hogy maximalizáljuk az általános lineáris modell (GLM) regresszor függetlenségét a feltételek között és optimalizáljuk a tervezési hatékonyságot (Buracas és Boynton, 2002). A résztvevők nem tudták megjósolni, hogy egy jelzőt egy inger követ-e vagy sem.
Az egyes próbák időtartama 4500 ms volt. A jelzőket t = 0-nál 1500 ms-ig, az ingereket pedig t = 2000-nél, szintén 1500 ms-ig mutatták be (1. ábra). A feladatot két, 68 próbából álló sorozatban hajtottuk végre. Az alapvonal három 30 s-os pihenőidőszakból állt, a két futás előtt, után és a két futás között. A feladat teljes időtartama 11,7 perc volt. Ez volt a munkamenet második feladata, és azután kezdődött, hogy a résztvevők körülbelül 10 percet feküdtek a szkennerben.
A résztvevőket arra utasították, hogy a jelző információi alapján készüljenek fel egy könnyű vagy nehéz ingerre (1. ábra), és a lehető legpontosabban és leggyorsabban jelezzék az ingerben lévő nyilak többségének irányát. A résztvevők a szkenneren kívül gyakorolták a feladatot egy 3 perces standard gyakorló protokollt követve. Ezt a protokollt megismételték, ha a teljesítmény nem volt elégséges (70% alatti pontosság a nehéz ingereknél).
A fejtekercshez rögzített tükör lehetővé tette a résztvevők számára, hogy lássák a fej mögött elhelyezett átlátszó vetítővásznat. A szkennerteremben lévő videoprojektor vetítette a feladat ingereit a képernyőre. A válaszok rögzítésére két pneumatikus nyomógombos, légnyomásos doboz szolgált. A résztvevők a bal hüvelyk- vagy mutatóujjukat használták a “bal” válaszhoz, a jobbat pedig a “jobb” válaszhoz.”
Képek felvétele
A felvételeket egy 3T Philips Achieva szkenneren (Philips Medical Systems, Best, Hollandia) végezték 32 csatornás SENSE fejtekercs használatával. Az anatómiai regisztrációhoz 3D T1-súlyozott szerkezeti képet szereztünk . fMRI-képeket 3D PRESTO impulzusszekvenciával nyertünk . A szkenner hat álfelvételt készített, majd elvetette őket.
Képek előfeldolgozása
Az fMRI-adatokat statisztikai parametrikus leképezéssel (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, London, UK2) előfeldolgozták. Az egyik ülésből származó szkenneléseket az első szkenneléshez igazítottuk, hogy korrigáljuk az alany mozgását a legkisebb négyzetek megközelítésével, hat paraméteres (merev test) térbeli transzformációval és 2. fokú B-spline becsléssel. A szkenneléseket egy merevtest-modell segítségével a T1-hez társregisztrálták. A paramétereket a normalizált kölcsönös információfüggvény segítségével becsültük. A képeket egy 4. fokú B-spline segítségével szeleteltük újra. A T1-et térbelileg normalizáltuk a standard MNI-térbe nagyon enyhe torzítás-regulázás (0,0001) és egy 4. fokú B-Spline segítségével. Az így kapott paramétereket az összes funkcionális felvételre alkalmaztuk az anatómiai különbségek minimalizálása és ezáltal a csoportos elemzés lehetővé tétele érdekében. Végül minden szkennelést térbelileg simítottunk egy 3D Gauss-szűrővel (teljes szélesség félmaximumon: 8 mm), hogy tovább minimalizáljuk a funkcionális anatómiai különbségek hatását.
Egyéni fMRI-elemzés
Egyéni GLM regressziós elemzést végeztünk minden egyes voxelre az egyéni aktivációs térképek létrehozásához egy tömeg-univariáns megközelítéssel, globális közelítő AR(1) autokorrelációs modellel és egy magaspass szűrővel (128 s cut-off). Az alapvonalat nem modelleztük explicit módon. Az alapfüggvény egy kanonikus HRF volt deriváltak nélkül. A maszkolási küszöbérték 0,8 volt. Minden feltételhez külön regresszorokat használtunk (CueE, 17 időpont; CueD, 16 időpont; StimE, 17 időpont; StimD, 18 időpont; és egy zavaró regresszor a feladatban az m-szekvenciához kapcsolódó blanc időszakokra, 68 időpont). Egyetlen időpontot sem zártunk ki az elemzésből. A béta-térképeket transzformáltuk, hogy tükrözzék a tényleges százalékos jelváltozást az egyes voxelekben. Csak a CueE és CueD regresszorokra vonatkozó eredményeket mutatjuk be, amelyek az inger nélküli jelváltozásokat tükrözik. A béta és a statisztikai t-térképeket vizuálisan ellenőriztük a nagyobb artefaktumok szempontjából.
Group fMRI Analysis
A második szintű fMRI-elemzést lineáris regresszióval végeztük minden egyes voxelen a vizualizáció és az érdeklődési terület (ROI) kiválasztása érdekében. A ROI-elemzést a GNU adatnyelvvel (GDL3) végeztük el, a GLM-elemzés által generált egyéni alanyi százalékos jelváltozási térképek felhasználásával. A ROI-n vagy hálózaton belüli voxeleket átlagoltuk a CueE, CueD és az alapvonal közötti jelkülönbség kiszámításához.
ROI kiválasztás
A CueE és a CueD esetében is SPM segítségével határoztuk meg a szupratentoriális helyi maximumokat és minimumokat korrigálatlan küszöbértékkel (p < 0,001; 1. táblázat). Az előre meghatározott méretű és alakú kubikus ROI-kat a CueE-alapvonal és a CueD-alapvonal kontraszt lokális maximumai fölé helyeztük külön-külön egy előre meghatározott raszteren belül (Jansma és Rutten, 2017; 2. táblázat). A ROI-k akkor kerültek be az elemzésbe, ha szignifikáns helyi maximumot tartalmaztak a CueE vagy a CueD kontrasztra vonatkozóan. Azzal, hogy a ROI-kat mindkét feltételhez külön-külön határozzuk meg a lokális maximumokon, megakadályozzuk a torzítást a CueE vagy a CueD felé. Továbbá a ROI-k előre meghatározott alakja és mérete a raszteren belül csökkenti a körkörösség hatását, mivel ezen ROI-k határait nem befolyásolja a zaj (Kriegeskorte et al., 2009). Továbbá ez a módszer megkönnyíti az eredmények kvantitatív összehasonlítását a különböző körülmények között, és potenciálisan a különböző vizsgálatok között, ezáltal elősegítve az fMRI-eredmények kvantitatív reprodukálhatóságát. A ROI-k elhelyezése a lokális maximumok felett optimalizált teljesítmény. Az aktivitáscsúcs helyét azonban köztudottan befolyásolja a zaj, ezért a ROI-k pontos helye mindig reprodukálható az egyes ROI-k esetében. Ez nem befolyásolja a hálózati eredményeket, mivel az összes ROI-t átlagoljuk, és a pontos elhelyezkedés ezért nem annyira releváns.
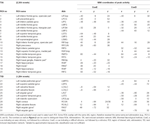
Táblázat. Tevékenységi minta.
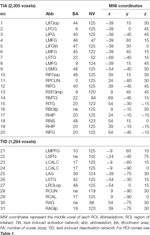
2. táblázat. Az érdeklődési terület (ROI) jellemzői.
A jelváltozás növekedését mutató régiók feletti ROI-kat egy feladat által kiváltott aktivációs hálózatban (“TIA”) kombináltuk. A jelváltozás csökkenését mutató régiók feletti ROI-kat a feladat által kiváltott deaktivációs hálózatban (“TID”; 1., 2. táblázat) kombináltuk. Ahol lehetséges volt, a ROI-k nevét az AAL atlasz (Tzourio-Mazoyer et al., 2002) segítségével határoztuk meg.
A 2. táblázat és a 2. ábra áttekintést nyújt az összes ROI méretéről és elhelyezkedéséről. Mivel a ROI-k mérete és határai előre meghatározottak, lehetséges, hogy az átlagos aktivitás a ROI-ban nem különbözik jelentősen az alapértéktől, például azért, mert az aktivált voxelek száma a ROI-n belül viszonylag kicsi. Bár a hálózati szintű átlagos aktivitás minden feltétel esetében szignifikánsan különbözik az alapértéktől (3. táblázat), a CueE esetében a 31 egyedi ROI-ból nyolc, a CueD esetében pedig a 31 ROI-ból három esetében nem ez a helyzet (4. táblázat). Csak olyan voxeleket vontunk be az elemzésbe, amelyek minden résztvevő esetében jelet tartalmaztak. Egy ROI maximális mérete 125 voxel, egyes ROI-k 125 voxelnél kevesebbet tartalmaznak, mert a koponya közelében helyezkednek el.
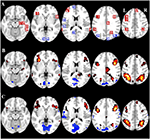
2. ábra. Az érdeklődési régió (ROI) elhelyezkedésének és a teljes agyi fMRI-eredmények áttekintése. (A) ROI-k a feladat által kiváltott aktivációhoz (piros) és a feladat által kiváltott deaktivációhoz (kék) Megjegyzés: a 22-es ROI nem jelenik meg, mivel a hely a kiválasztott szeleteken kívül esik. A ROI-k MNI-koordinátái, BA-száma és mérete a 2. táblázatban található. (B) A CueE t-értékeinek vizuális ábrázolása az alapvonalhoz képest (≥3 = pirosból sárgába, ≤ -3 = kékből világoskékbe), (C) a CueD t-értékei az alapvonalhoz képest (≥3 = pirosból sárgába, ≤ -3 = kékből világoskékbe). A képek neurológiai orientációban vannak (L = balra, R = jobbra), a csúcsaktivációs voxelek nevei, rövidítései, MNI-koordinátái az 1. táblázatban találhatók.

3. táblázat. Hálózati eredmények.
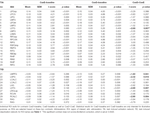
Táblázat. ROI eredmények.
hipotézisek tesztelése
A következő hipotéziseket teszteltük ebben a vizsgálatban:
1. hipotézis: A TIA a CueD esetében fokozott jelváltozást mutat a CueE-hez képest.
2. hipotézis: A TID csökkent jelváltozást fog mutatni a CueD esetében, a CueE-hez képest.
Statisztikai elemzés
Teljesítményelemzést nem végeztünk, mivel ez volt az első alkalom, hogy ezt a speciális feladatkialakítást bemutattuk a résztvevőknek. Ezért nincs ismeretünk a körülmények várható százalékos jelváltozásáról és variabilitásáról (Desmond és Glover, 2002).
A feladat pontosságát a StimE és a StimD esetében a helyes válaszok százalékos arányaként számoltuk ki. A reakcióidőt a StimE és a StimD esetében az összes helyes válaszra vonatkozóan számoltuk ki.
Egymintás t-próbát alkalmaztunk a TIA és a TID százalékos jelváltozásának összehasonlítására a CueE és a CueD esetében a kiindulási értékhez képest. Páros mintás egyoldalú t-próbákat használtunk a CueD és a CueE közötti jelváltozás, pontosság és reakcióidő különbségek összehasonlítására (1. és 2. hipotézis).
A post hoc tesztelés során az egyes hálózatokon belül minden egyes ROI-ra külön t-próbákat végeztünk. A ROI-elemzés Bonferroni-korrigált volt, ha a hálózati omnibusz teszt nem volt szignifikáns. Bemutatjuk a nem korrigált p-értékeket és egy megjegyzést, amely szerint ez a p-érték minden többszörös összehasonlítás esetén túlélte volna a Bonferroni korrekciót. Minden statisztikai elemzéshez az SPSS 24 programot használtuk.
Eredmények
Teljesítmény
A könnyű ingerek (“StimE”) pontossága 98% ± 1,2% SEM, a nehéz ingerek (“StimD”) 85% ± 2,6% volt. A reakcióidő a “StimE” esetében 682 ± 20 ms, a “StimD” esetében 1.138 ± 27 ms volt. Mindkét feltétel elegendő pontossági arányt mutat ahhoz, hogy biztosak lehessünk abban, hogy a feladatot minden résztvevő az utasításoknak megfelelően hajtotta végre. Ahogy feltételeztük, a résztvevők a StimD esetében kevésbé pontosan (T = -4,3, p < 0,001) és lassabban (T = 17,8, p < 0,001) reagáltak, mint a StimE esetében (3. ábra), ami azt jelzi, hogy a könnyű és nehéz ingerek közötti kontraszt sikeres volt.
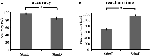
3. ábra. Teljesítményeredmények. (A) Pontosság mindkét inger-kategóriára, (B) reakcióidő mindkét inger-kategóriára. A hibasávok az átlag standard hibáját (SEM) jelentik. *p ≤ 0.001. Rövidítések: StimE, könnyű ingerre utaló jelzés, amelyet könnyű inger követ; StimD, nehéz ingerre utaló jelzés, amelyet nehéz inger követ.
Funkcionális MRI
Descriptives
A TIA-hálózat 20 ROI-ból állt, amelyek közül 10 a CueE helyi maximumain alapult, öt a CueD helyi maximumain alapult, és öt tartalmazott egy CueE és egy CueD maximumot is (1. táblázat). Ebből a 20 ROI-ból 19 mutatott szignifikáns aktivitást a kiindulási értékhez képest a CueE esetében, 18 a CueD esetében (1. táblázat). A TIA átlagos jelváltozása a kiindulási értékhez képest a CueE esetében 0,12 ± 0,02 (t = 6,11, p < 0,001), a CueD esetében 0,12 ± 0,02 (t = 7,61, p < 0,001) volt.
A TID-hálózat 11 ROI-ból állt, amelyek közül három a CueE helyi maximumain alapult, hat a CueD helyi maximumain alapult, és kettő tartalmazott egy CueE és egy CueD maximumot is (1. táblázat). Ebből a 11 ROI-ból négy mutatott szignifikáns aktivitást a kiindulási értékhez képest a CueE esetében, 10 pedig a CueD esetében (1. táblázat). A TID átlagos jelváltozása a kiindulási értékhez képest a CueE esetében -0,07 ± 0,02 (t = 2,68, p < 0,01), a CueD esetében -0,12 ± 0,02 (t = 5,37, p < 0,001; 3. táblázat).
A várt nehézséghez való hangolás
A TIA jelnövekedése nem volt szignifikánsan erősebb a CueD esetében a CueE-hez képest (t = -0,24, p = 0,41), ami azt jelzi, hogy a TIA aktiváció szintje nem hangolódik a várt nehézséghez (4. ábra, 3. táblázat). A post hoc ROI-elemzés a Bonferroni korrekció alkalmazása után a TIA-n belüli 20 ROI egyike sem mutatott szignifikáns jelnövekedést a CueD esetében a CueE-hez képest (4. táblázat, 5. ábra).
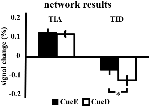
4. ábra. A hálózati eredmények áttekintése. A hibasávok SEM értéket jelentenek. *p ≤ 0,05 a nehéz vs. könnyű jelszó esetében. Rövidítések: TIA, feladat indukálta aktivációs hálózat; TID, feladat indukálta deaktivációs hálózat; CueE, könnyű ingerre utaló jelző, amelyet nem követ inger; CueD, nehéz ingerre utaló jelző, amelyet nem követ inger.
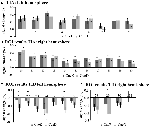
5. ábra. Az egyes ROI-válaszok áttekintése a feladat által kiváltott aktivációs hálózaton belül, bal (A) és jobb félteke (B), valamint a feladat által kiváltott deaktiváció, bal (C) és jobb félteke (D). A hibasávok SEM-et jelentenek. *p ≤ 0,05 a CueD vs. CueE esetében, a ROI-k elhelyezkedését lásd a 2. ábrán, a ROI-k nevét, rövidítéseit és MNI koordinátáit lásd az 1. táblázatban.
A TID jelcsökkenése szignifikánsan erősebb volt a CueD esetében a CueE-hez képest (T = -2,48, p = 0,01), ami azt jelzi, hogy a TID szintje az elvárt nehézséghez van hangolva (3. táblázat, 4. ábra). A post hoc ROI-elemzés a TID 11 ROI-jából négyben mutatott szignifikáns jelcsökkenést a CueD esetében a CueE-hez képest, nevezetesen az LMPFG (T = -1,83, p = 0,04), LSPG (T = -2,32, p = 0,02), LSTG (T = -2,69, p < 0,01) és RCALC (T = -2,90, p < 0,01). Ezek az eredmények azt mutatják, hogy a várható nehézségre való hangolás a legerősebb a bal frontális, temporális és vizuális ROI-kban (4. táblázat, 5. ábra).
Diszkusszió
Ebben a tanulmányban azt vizsgáltuk, hogy a fázisos éberséget a közelgő kognitív kihívás várható nehézségére hangolják-e és hogyan. A fázisos éberség tükröződött az aktivált és a deaktivált agyi régiókban egyaránt. Az aktivált hálózatba a központi végrehajtó hálózat régiói tartoztak. A deaktivált hálózat a DMN régióit, valamint a vizuális agykérget foglalta magában. A deaktiváció szintje a közelgő inger várható nehézségével skálázódott, míg az aktiváció szintje nem. Ezek az eredmények arra utalnak, hogy a fázisos éberség modulációja túlnyomórészt az aktuális feladat szempontjából irreleváns, fokozatos szuppressziós folyamatokban (például kardio-vagális, auditív és vizuális folyamatokban) tükröződik. Ez valószínűleg azért következett be, mert ezek a folyamatok zavarhatják a közelgő kognitív kihívásokat.
A fázisos éberséggel kapcsolatos korábbi képalkotó vizsgálatok egy nehézségi szintet használtak, csak aktivált agyi régiókat mutattak be (Shulman et al., 1999; Sturm és Willmes, 2001; Thiel et al., 2004; Fan et al., 2005; Périn et al., 2010; Yanaka et al., 2010), vagy kizárólag a vizuális kéreg aktivitására összpontosítottak (Bartolucci és Smith, 2011). Ezek a vizsgálatok egy többnyire jobb oldali-laterális fronto-parietális-thalamikus hálózatot azonosítottak, néha motoros és vizuális agykérgi régiókkal kombinálva, amelyet egy jelző aktivált. A tanulmányok között jelentős különbségek vannak az aktivált hálózatokban. A korábbi tanulmányok hálózatai közötti különbségek egy része a jelzést követő feladattal magyarázható. Például Thiel és munkatársai (2004) csak a vizuális agykérgi régiókban azonosítottak aktivációt vizuális észlelési feladattal, míg Fan és munkatársai (2005) és Yanaka és munkatársai (2010) talamikus és motoros aktivációt azonosítottak motoros válaszfeladattal. Bár Bartolucci és Smith (2011) egy elegáns cued task designt mutatott be egy orientációs diszkriminációs feladat négy nehézségi szintjével, ők csak a vizuális kéregben mutattak be aktivációt. Ezért nem világos, hogy más agyi régiók hogyan reagáltak ebben a feladatban. A feladat által kiváltott aktivációs hálózat, amelyet a jelenlegi vizsgálatban azonosítottak, leginkább hasonlít a Shulman és munkatársai (1999) által talált hálózatra, amely hasonló típusú vizuális feldolgozási feladatot használt.
Nem világos, hogy a fázisos éberség alulról felfelé vagy felülről lefelé irányuló válasz-e egy nem informatív jelre (Thiel és munkatársai, 2004; Hackley, 2009; Périn és munkatársai, 2010; Bartolucci és Smith, 2011; Chica és munkatársai, 2016). A fázisos éberséget egyesek bottom-up (Sturm et al., 1999; Hackley, 2009), mások top-down (Bartolucci és Smith, 2011), megint mások pedig kombinált (Thiel et al., 2004; Périn et al., 2010) válasznak tekintik. Azt állítjuk, hogy a legmegfelelőbb a jelzőre adott agyi választ az alulról felfelé és felülről lefelé történő feldolgozás kombinációjaként címkézni, mivel a résztvevők mind a külső jelzőre, mind az ehhez a jelzőhöz kapcsolódó utasításra reagálnak. Fő eredményeink azonban a két hasonló, különböző típusú feladatserkentésre utaló jelzésekre adott válaszok különbségére vonatkoznak. Ezért azt állítjuk, hogy a nehéz feladatot jelző jelző és a könnyű feladatot jelző jelző közötti eredő különbség a top-down kontroll egy olyan szintjét képviselheti, amely a jelző és a kapcsolódó utasítás értelmezéséhez kapcsolódik.
A feladat által indukált deaktivációs hálózat a DMN agyi régióit foglalta magában, nevezetesen a mediális prefrontális gyrus, a felső temporális gyri és a szögletes gyri (Shulman et al., 1999; Raichle et al., 2001; Raichle, 2015). Általánosan feltételezik, hogy a DMN deaktiválása segíti a feladatteljesítményt azáltal, hogy csökkenti a kognitív kihívásokat zavaró belső folyamatokat (Raichle et al., 2001; Buckner et al., 2008; Raichle, 2015). Ezt a nézetet alátámasztják azok a vizsgálatok, amelyek azt mutatják, hogy a DMN deaktiváció szintje a feladat végrehajtása során a feladat nehézségével skálázódik (McKiernan et al., 2003; Singh és Fawcett, 2008). Ezek a tanulmányok a feladat végrehajtása során vizsgálták a deaktivációt. Eredményeink arra utalnak, hogy ez a skálázás már a feladat anticipációs fázisában is bekövetkezhet. Weissman és munkatársai (2006) az egyéni reakcióidők változékonyságát vizsgálták a figyelem kiesésének azonosítására. A figyelem kiesései összefüggésbe hozhatók a DMN csökkent deaktivációjával. Míg (Weissman és munkatársai, 2006) a figyelem szintjének természetes variációját vizsgálták, addig a mi eredményeink azt mutatják, hogy a feladat ingerének bemutatása előtt több DMN régió deaktivációja is hangolható top-down módon.
A legmélyebb hangolási hatásokat a bal oldali mediális prefrontális, temporális és vizuális régiókban találtuk. Az alábbiakban e hatások lehetséges következményeit tárgyaljuk ezekre a régiókra vonatkozóan a javasolt funkciójukkal kapcsolatban.
A mediális prefrontális kéreg aktivitását korábban összefüggésbe hozták az elme vándorlásával (Bertossi et al., 2017), az emlékezet-visszakereséssel (Euston et al., 2012) és a kardiovagális kontrollal (Wong et al., 2007). Ez a mediális prefrontális deaktiváció arra utal, hogy ezen funkciók bármelyikét már a kognitív kihívást megelőzve elnyomják.
A deaktiváció hangolása a várható nehézséghez a vizuális és temporális régiókban összefügghet az irreleváns vizuális és auditív szenzoros bemenet elnyomásával. A kétoldali temporális régiókban történő hangolás kifejezetten az MRI-szkenner által keltett zaj elnyomásához kapcsolódhat. Ésszerűen elvárható, hogy a zajszint hasonló legyen a körülmények között. Az a tény, hogy a deaktiváció szintje erősebb a nehéz jelzések esetében, arra utal, hogy a környezeti hangok feldolgozása erősebben elnyomódik, ha nehéz feladat várható. A vizuális agykérgekben történő deaktivációról ritkábban számolnak be, mint a temporális régiókban történő deaktivációról. Smith és munkatársai (2000) azonban korábban összefüggésről számoltak be a vizuális kéregben elterjedt deaktiváció és a kép egy bizonyos részére irányuló figyelem között. Giesbrecht és munkatársai (2006) szintén beszámoltak a vizuális aktiváció modulációjáról, amely a látómező középpontjához közeli tárgyakra irányuló rejtett figyelemhez kapcsolódik a perifériás látómezőben lévő tárgyakhoz képest. Ezek az eredmények azt jelzik, hogy a vizuális agykérgekben az aktiváció modulálható a foveális vagy a perifériás területekre irányuló rejtett figyelem által. Vizsgálatunk új dimenzióval egészíti ki ezeket az eredményeket, mivel azt sugallja, hogy a vizuális agykérgekben az aktivációt nemcsak a rejtett vizuális figyelem helye modulálja, hanem a vizuális kihívás várható nehézsége is; és mivel a jelzések helyét és méretét tekintve azonosak voltak, úgy tűnik, hogy ez a folyamat felülről lefelé szabályozott.
Egy sor olyan agyi régiót is azonosítottunk, amelyek fokozott aktivációt mutattak egy kognitív kihívás előrejelzésekor. Ez a hálózat hasonlóságot mutatott a központi végrehajtó hálózattal, amelyet korábban a munkamemória feladatok végrehajtásával hoztak összefüggésbe (Lawrence et al., 2003). Ennek a hálózatnak az aktivációja azonban nem volt a várható feladat nehézségére hangolva. Ez arra utal, hogy ezek a régiók olyan folyamatokat képviselnek, amelyek hasonlóak a nehéz és a könnyű feltétel esetében. Lehetséges, hogy az aktivitás ebben a hálózatban a jelző értékeléséhez, vagy a feladatkontextus és az utasítások előhívásához és fenntartásához kapcsolódik.
Meg kell fontolni néhány korlátozást, amelyek befolyásolhatják eredményeink értelmezését. Először is, nem vontunk be olyan próbákat, amelyekben az ingereket nem előzte meg súgó, ezért nem lehet vizsgálni a súgó hatását a kognitív teljesítményre. Az idő szűkössége miatt nem volt lehetőségünk megismételni a feladatunkat azonos ingerekkel, de jelzések nélkül.
Második, a nehéz kognitív kihívást jelző jelző részeként felkiáltójelet használtunk, mivel ezt a szimbólumot jellemzően akkor használják, ha fokozott éberségre van szükség. Lehetséges, hogy a felkiáltójel maga is szerepet játszott a könnyű és a nehéz feltétel eltérő aktivációs mintázatában, a benne rejlő jelentése miatt. Ez azonban nem befolyásolja eredményeink fő értelmezését, kivéve, hogy fő eredményeinket a felülről lefelé irányuló válasz helyett alulról felfelé irányuló válaszként lehet besorolni. Végül, kísérleti elrendezésünk nem teszi lehetővé annak vizsgálatát, hogy a fázisos éberséggel kapcsolatos aktivitás általánosítható-e más tartományokra, vagy pedig tartományspecifikus, mivel csak vizuo-perceptuális kognitív kihívást tartalmazott.
Végezetül, ebben a tanulmányban kimutattuk, hogy a fázisos éberség hangolását több régióban a deaktiváció szintje reprezentálja. Ez a hatás a mediális prefrontális, vizuális és temporális kéregben a legerősebb, ami a kardiovagális kontroll elnyomását, a perifériás látómező vizuális feldolgozását és az MRI-szkenner zajának elnyomását tükrözheti. Ezek az eredmények arra utalnak, hogy a kognitív teljesítményt elősegíti az agy olyan állapota, amely szorosan kapcsolódik a közelgő kognitív kihívás nehézségére vonatkozó várakozásokhoz.
Adatok elérhetősége
A nyers adatok az Elisabeth-Tweesteden kórházban keletkeztek. A tanulmány megállapításait alátámasztó származtatott adatok ésszerű kérésre elérhetők a megfelelő szerzőtől.
Etikai nyilatkozat
Ezt a tanulmányt a holland szociális támogatási törvény ajánlásainak megfelelően végeztük, orvosi etikai bizottság Brabant. Minden alany a Helsinki Nyilatkozatnak megfelelően írásbeli tájékozott beleegyezését adta. A protokollt a Brabant orvosi etikai bizottsága hagyta jóvá.
A szerzők hozzájárulása
MD és JJ írta a fő kéziratot és készítette az ábrákat. MD és IS végezte a kísérleteket. G-JR, NR és JJ közösen felügyelték a munkát. Minden szerző átnézte a kéziratot.
Finanszírozás
Ezt a tanulmányt a ZonMW, egy holland nemzeti szervezet (Netherlands Organization for Health Research and Development, ZONMW; Project number: 842003004) finanszírozta, a projekt címe:
Érdekütközéssel kapcsolatos nyilatkozat
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségként értelmezhetők.
Köszönet
Megköszönjük William Pigmans és Maikel Brands radiográfusoknak a tanulmányhoz való hozzájárulásukat.
Rövidítések
DMN, default mode network; M, férfi; F, nő; GLM, általános lineáris modell; SPM, statisztikai parametrikus leképezés; CueE, könnyű ingerre utaló jel, amelyet nem követ inger; CueD, nehéz ingerre utaló jel, amelyet nem követ inger; StimE, könnyű ingerre utaló jelző, amelyet könnyű inger követ; StimD, nehéz ingerre utaló jelző, amelyet nehéz inger követ; MRI, mágneses rezonancia képalkotás; ROI, region of interest; GDL, GNU data language; TIA, task induced activation network; TID, task induced deactivation network; SEM, standard error of mean.
Lábjegyzetek
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M., and Smith, A. T. (2011). A figyelemmoduláció a vizuális kéregben módosul a perceptuális tanulás során. Neuropsychologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi, E., Peccenini, L., Solmi, A., Avenanti, A., and Ciaramelli, E. (2017). A mediális prefrontális kéreg transzkraniális egyenáramú stimulációja tompítja az elme vándorlását férfiaknál. Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., and Schacter, D. L. (2008). Az agy alapértelmezett hálózata: anatómia, funkció és jelentősége a betegségek szempontjából. Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Buracas, G. T., and Boynton, G. M. (2002). Eseményfüggő fMRI-kísérletek hatékony tervezése M-szekvenciák segítségével. Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | CrossRef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P., and Bushnell, M. C. (2015). Szükség van-e egy reszponzív alapértelmezett üzemmódú hálózatra a sikeres munkamemóriafeladat-teljesítményhez? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., and Paz-Alonso, P. M. (2016). A fázisos riasztás és a tudatosság közötti kölcsönhatások a fronto-striatális hálózatban. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | CrossRef Full Text | Google Scholar
Desmond, J. E., and Glover, G. H. (2002). A mintanagyság becslése funkcionális MRI (fMRI) neuroimaging vizsgálatokban: statisztikai teljesítményelemzések. J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., and McNaughton, B. L. (2012). A mediális prefrontális kéreg szerepe a memóriában és a döntéshozatalban. Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., and Posner, M. I. (2005). A figyelmi hálózatok aktiválása. Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., and Posner, M. I. (2002). A figyelmi hálózatok hatékonyságának és függetlenségének tesztelése. J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/089892902317361886
PubMed Abstract | CrossRef Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G., and Mangun, G. R. (2006). A célpont előtti aktivitás a vizuális kéregben előre jelzi a viselkedési teljesítményt a térbeli és jellemző figyelemfeladatokban. Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009). Az önkéntes reakció felgyorsítása figyelmeztető jelzéssel. Psychophysiology 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R., Sorsa, M., and Siivonen, P. (2008). A 6/6-os és 4/8-as őrszolgálati rendszerek hatása a hídőrök álmosságára. Chronobiol. Int. 25, 413-423. doi: 10.1080/07420520802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A., and Buckner, R. L. (2012). A figyelemszabályozás modulációjának kudarca az előrehaladott öregedés során a fehérállomány patológiájához kapcsolódik. Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., and Duyn, J. H. (2007). fMRI study of effort and information processing in a working memory task. Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., and Rutten, G. J. (2017). “A proposal for a system that facilitates quantitative replication of fMRI studies,” in Proceedings of the Human Brain Mapping Conference 2017 June 25-29, Vancouver, Canada. Online elérhető a következő címen: https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., and Baker, C. I. (2009). Körkörös elemzés a rendszeridegtudományban: a kettős merítés veszélyei. Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A., and Bartolomeo, P. (2011). A fázisos auditív riasztás javítja a vizuális tudatos észlelést. Conscious. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G., and Stein, E. S. (2003). Több neurális hálózat közvetíti a fenntartott figyelmet. J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/089892903770007416
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, G., Sobering, G., Duyn, J., and Moonen, C. T. (1993). A funkcionális MRI technika, amely a visszhangeltolódás elveit kombinálja egy megfigyeléssorozattal (PRESTO). Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | CrossRef Full Text | Google Scholar
Macleod, J. W., Lawrence, M. A., McConnell, M. M., Eskes, G. A., Klein, R. M., and Shore, D. I. (2010). Az ANT értékelése: a figyelemhálózati teszt pszichometriai és elméleti megfontolásai. Neuropsychology 24, 637-651. doi: 10.1037/a0019803
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., D’Angelo, B. R. D., Kaufman, J. N., and Binder, J. R. (2006). A “tudatfolyam” megszakítása: egy fMRI-vizsgálat. Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J. K., and Binder, J. R. (2003). A feladat által kiváltott deaktivációt befolyásoló tényezők parametrikus manipulációja a funkcionális neuroimagingben. J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/089892903321593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R. D., and Brady, C. B. (1993). Fázisos és tónusos éberség az Alzheimer-kórban. Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., and Ramsey, N. F. (2008). Fokozott érzékenység gyors háromdimenziós vér-oxigénszint-függő funkcionális MRI-vel: a SENSE-PRESTO és a 2D-EPI összehasonlítása 3 T-nél. NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., and de Marco, G. (2010). Éberség fiatal egészséges alanyoknál: a figyelmeztető jel által fokozott agyi régiók interaktivitásának fMRI-vizsgálata. Brain Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H., et al. (2009). A munkamemória terhelés hatása az FMRI nyugalmi állapot mintázatára a későbbi nyugalmi fázisokban. PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E. (2015). Az agy alapértelmezett üzemmódú hálózata. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E., MacLeod, J. W., Snyder, A. Z., Powers, W. J., Gusnard, D. A., and Shulman, G. L. (2001). Az agyműködés alapértelmezett módja. Proc. Natl. Acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Petersen, S. E., et al. (1999). A mozgó tárgyakra vonatkozó irányelvárások kódolásában és alkalmazásában részt vevő területek. J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K. D., and Fawcett, I. P. (2008). Az emberi alapértelmezett üzemmódú hálózat átmeneti és lineárisan fokozatos deaktiválása vizuális észlelési feladattal. Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, A. T., Singh, K. D., and Greenlee, M. W. (2000). Az emberi látókéreg aktivitásának figyelmi elnyomása. Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., de Simone, A., Krause, B. J., Specht, K., Hessemann, V., Radermacher, I., et al. (1999). Az intrinzikális éberség funkcionális anatómiája: bizonyíték egy froto-parietális-thalamikus-agytörzsi hálózatra a jobb féltekében. Neuropsychologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., and Willmes, K. (2001). Az intrinzik és a fázisos éberség funkcionális neuroanatómiájáról. Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | CrossRef Full Text | Google Scholar
Thiel, C. M., Zilles, K., and Fink, G. R. (2004). A vizuospatialis figyelem éberségének, tájékozódásának és átirányításának agyi korrelátumai: egy eseményhez kapcsolódó fMRI vizsgálat. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Az aktivációk automatizált anatómiai jelölése SPM-ben az MNI MRI egyszemélyes agyának makroszkopikus anatómiai parcellázásával. Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | CrossRef Full Text | Google Scholar
van Gelderen, P., Ramsey, N. F., Liu, G., Duyn, J. H., Frank, J. A., Weinberger, D. R., et al. (1995). Az emberi agy háromdimenziós funkcionális mágneses rezonancia képalkotása klinikai 1,5-T szkenneren. Proc. Natl. Acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N., and Henik, A. (2011). A fázisos éberség a vizuális ingerek globális feldolgozásának fokozásával modulálhatja a végrehajtó kontrollt. Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., Visscher, K. M., and Woldorff, M. G. (2006). A figyelem pillanatnyi kihagyásainak idegi alapjai. Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | CrossRef Full Text | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S., and Shoemaker, J. K. (2007). Ventrális mediális prefrontális kéreg és a kardiovagális kontroll tudatos emberekben. Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka, H. T., Saito, D. N., Uchiyama, Y., and Sadato, N. (2010). A fázisos éberség idegi szubsztrátjai: funkcionális mágneses rezonancia képalkotó vizsgálat. Neurosci. Res. 68, 51-58. doi: 10.1016/j.neures.2010.05.005
PubMed Abstract | CrossRef Full Text | Google Scholar