Introduction
これまでの研究で、人は手がかりを利用して認知課題に精神的に備えることができることが明らかにされています。 手がかりが提示された瞬間に精神的な準備をするよう被験者に指示した場合、これは位相的覚醒と定義されている。 覚醒度に影響を与える能力は重要であり、特に覚醒度にコストがかかる場合は重要である。 覚醒への要求が高すぎる場合、疲労(Härmä et al.2008)やストレス関連の問題につながる可能性がある。 本研究では、健康な成人が、今後予想される認知課題の難易度に合わせて覚醒度を調整するか、またどのように調整するかを調べることを目的とする。
課題刺激の直前に手がかりが提示されると、一般に人は単純な運動反応課題で速く反応でき(ファンら、2002;マクラウドら、2010;ワインバックとヘニック、2011)、劣化した視覚刺激をより正確に知覚できる(クスニァら、2011)。 画像研究により、位相性覚醒は前頭葉、頭頂葉、視床(Shulmanら、1999;Fanら、2005;Yanakaら、2010)、側頭葉(Thielら、2004)、視覚脳領域(Bartolucci and Smith、2011)の活性化の増加と関連することが示されています。 これらの画像研究により、脳内における位相的覚醒の表現について貴重な情報が得られている。 しかし、これまで位相性覚醒は一般にオン/オフ現象として研究されてきたか、あるいは解析対象が視覚野に限定されてきました。 さらに、これまでの研究で、デフォルトモードネットワーク(DMN)の一部である脳領域の不活性化が、認知課題の遂行を促進する可能性があることが示されている。 まず、タスク実行に関する研究では、タスクの難易度の上昇は、不活性化の上昇と関連していることが示されている(McKiernanら、2003、2006;Jansmaら、2007;Singh and Fawcett、2008;Pykaら、2009;Heddenら、2012;Čekoら、2015)。 次に、Jansmaら(2007)は、DMNの内側前頭葉部分は、参加者が予測できる課題の難易度にのみ同調し、参加者が予測できない各刺激の実際の難易度には影響を受けないことを示しました。 最後に、Weissmanら(2006)は、被験者の反応が比較的遅い場合、刺激直前のDMNの不活性化が弱くなることを見出した。 3545>本研究では、簡単な条件と難しい条件からなる手がかり付き視覚認知課題を設計した。 手がかりはこれから来る刺激の難易度に関する情報を提供するが、課題そのものを容易にするような情報は提供しない。 半数の試行では、手がかりの後に刺激が与えられなかった。 これらの試行のみ脳活動を分析し、手がかりに関連する活動と課題遂行に関連する活動を完全に分離することができた。 我々は、参加者はタスクの予想される難易度に合わせて活動を調整していると仮定する。
Materials and Methods
Participants
参加者はオンライン広告で募集された。 参加者は,重大な神経疾患や精神疾患の既往がある場合,あるいは磁気共鳴画像法(MRI)検査の禁忌(体内または周囲の金属物,閉所恐怖症,妊娠)を報告した場合には除外された。 この研究は、人を対象とする医学研究法(WMO)、医学研究倫理委員会ブラバントの勧告に従って実施された。 プロトコルは、医学研究倫理委員会によって承認された(プロトコル番号:NL51147.028.14)。 すべての被験者はヘルシンキ宣言に基づき、書面によるインフォームドコンセントを行った。
22人の健康な右利きボランティアが研究に参加した。 2名の参加者のデータはスキャナーのアーチファクトにより除外された。
Task Design
我々は、タスクの実行による影響を交絡させずに、タスクの予想難易度に関連する予期効果を調べることができるタスク(事象関連デザイン)をデザインした。
タスク刺激は、黒い画面上に3×3のレイアウトで9つの矢印からなる(図1)。 簡単な刺激」は正しい方向の矢印が8個、反対方向の矢印が1個であった。 一方、「難しい刺激」は、正しい方向に5つ、反対方向に4つの矢印が配置されたものである(図1)。 参加者は、矢印の多い方向に対応する手でボタンを押すように指示された。 簡単な刺激と難しい刺激はランダムな順序で提示された。 ベースライン刺激は、テキストが表示された黒い静止画面から構成された。 ベースライン刺激は、オランダ語で「あなたは30秒の休憩があります」というテキストが書かれた黒いスクリーンで構成された。 刺激提示は “presentation “ソフトウェアによって行われた。 視覚刺激の表象。 各試行は4.5秒であり、半数の試行では手がかりの後に刺激がなかった。 これらの試行は機能的磁気共鳴画像法(fMRI)の解析に使用された。
各試行は、その後に提示される刺激の難度を示す合図で開始された。 緑色の点は「易しい刺激」、感嘆符付きの緑色の点は「難しい刺激」を示した(図1)。
試行の半分では手がかりの後に刺激がなく、残りの半分では手がかりが消えた500ミリ秒後に刺激が提示された。 この結果、4つの条件が成立した。 (1)簡単な刺激に対する手がかりの後に刺激を与えない(CueE),(2)難しい刺激に対する手がかりの後に刺激を与えない(CueD),(3)簡単な刺激に対する手がかりの後に簡単な刺激を与える( StimE),(4)difficult 刺激を与える( StimD)であった。 すべての条件はベースラインと対照された。 機能的MRI(fMRI)の結果は、CueEとCueDに基づくものであった。 パフォーマンス結果は、StimEとStimDに基づくものであった。 CueEとCueDは、条件間の一般線形モデル(GLM)回帰因子の独立性を最大化し、デザイン効率を最適化するために、Mシーケンスを用いて擬似ランダム化した順序で提示した(Buracas and Boynton, 2002)。 参加者は、手がかりの後に刺激が続くかどうかを予測することはできなかった。 図1)。合図はt = 0で1,500ms提示され、刺激はt = 2,000で同じく1,500ms提示された(図1)。 課題は68試行を2回に分けて行った。 ベースラインは、2回の試行の前、後、間に30秒の休息時間を3回設けた。 タスクの総時間は11.7分であった。 この課題はセッションの2番目の課題で、参加者がスキャナで約10分間横になってから開始された。
参加者は、キューの情報に基づいて易しい刺激または難しい刺激を準備し(図1)、刺激中の大部分の矢印の方向をできるだけ正確かつ迅速に指示するよう指示された。 参加者はスキャナの外で3分間の標準練習プロトコルにしたがって課題の練習を行った。 このプロトコルは、パフォーマンスが不十分な場合(難しい刺激に対して70%以下の精度)に繰り返された。
ヘッドコイルに取り付けられた鏡によって、参加者は頭の後ろに配置されたシースルーの投影スクリーンを見ることができた。 また、スキャナ室内に設置されたビデオプロジェクタにより、課題刺激がスクリーンに投影された。 反応の記録には空気圧式押しボタンボックス2個を使用した。 参加者は左手の親指または人差し指で「左」と答え、右手で「右」と答えた。
画像取得
スキャンは3T Philips Achieva scanner (Philips Medical Systems, Best, Netherlands) で32チャンネルのSENSEヘッドコイルを使用して行われた。 fMRI画像は3D PRESTOパルスシーケンスを用いて取得した。 3657>
画像前処理
fMRI データは statistical parametric mapping (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, London, UK2) を用いて前処理を施した。 1セッションのスキャンは、最小二乗法、6パラメータ(剛体)空間変換、2次Bスプライン推定を用いて、被験者の動きを補正するために最初のスキャンに再調整された。 スキャンは剛体モデルを用いてT1にコ・レジストレーションされた. パラメータは正規化相互情報関数を用いて推定した. 画像は4次B-splineによって再スライスされた. T1は非常に軽いバイアス正則化(0.0001)と4次B-スプラインを用いて標準的なMNI空間に空間的に正規化された. 解剖学的な差異を最小化し、グループ解析を可能にするために、得られたパラメータをすべての機能的スキャンに適用した。 最後に、すべてのスキャンを3Dガウスフィルタ(半値全幅:8mm)で空間的に平滑化し、さらに解剖学的な差異による影響を最小化した。
個別fMRI解析
事象関連GLM回帰分析を各ボクセルに対して行い、グローバルな近似AR(1)自己相関モデルとハイパスフィルタ(128秒カットオフ)による質量一変量アプローチで個別活性化図を作成した。 ベースラインは明示的にモデル化されなかった。 基本関数は微分を含まない正準HRFとした。 マスキング閾値は0.8を使用した。 各条件で別々のリグレッサーを用いた(CueE, 17 timepoints; CueD, 16 timepoints; StimE, 17 timepoints; StimD, 18 timepoints; およびm系列に関連するタスクのblanc期間の迷惑リグレッサー, 68 timepoints). 解析から除外されたタイムポイントはない。 ベータマップは、各ボクセルにおける実際の信号変化の割合を反映するように変換された。 CueEとCueDのリグレッサーのみ、刺激のないキューに対する信号変化を反映した結果を示す。 βマップと統計的tマップは、大きなアーチファクトがないか目視で確認した。
グループfMRI解析
第2レベルのfMRI解析は、可視化と関心領域(ROI)選択のために各ボクセルでの線形回帰で実施された。 ROI解析は、GLM解析によって生成された個々の被験者のパーセント信号変化マップを使用して、GNUデータ言語(GDL3)で実行されました。 ROIまたはネットワーク内のボクセルを平均し、CueE、CueD、ベースラインの信号差を算出した。
ROI Selection
SPMによりCueEとCueDについて、補正なしの閾値で局所最大値と最小値を求めた(p < 0.001; Table 1)。 あらかじめ定義されたラスター内のCueE-ベースラインおよびCueD-ベースラインのコントラストの局所最大値上に、あらかじめ定義されたサイズおよび形状の立方体ROIを別々に配置した(Jansma and Rutten, 2017; 表2)。 ROIは、CueEまたはCueDの有意な局所最大値を含む場合、分析に含まれた。 両条件の局所最大値に別々にROIを定義することで、CueEまたはCueDのどちらかに偏ることを防ぐことができます。 さらに、ラスター内のROIの形と大きさがあらかじめ定義されているため、これらのROIの境界はノイズの影響を受けないので、円形度の影響を軽減することができる(Kriegeskorte et al.) さらに、この方法は、条件間、潜在的には異なる研究間の結果の定量的比較を容易にするため、fMRI結果の定量的再現性を促進する。 局所最大値上にROIを配置することで、パワーを最適化することができた。 しかし、活動ピークの位置はノイズの影響を受けることが知られており、ROIの正確な位置はROIごとに常に再現される可能性があります。 すべてのROIは平均化され、正確な位置は、したがって、それほど関連していないので、これは、ネットワークの結果に影響を与えません。 活動パターン。
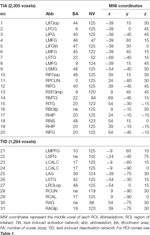
Table 2.活動パターン。 関心領域(ROI)の特徴。
信号変化が増加した領域上のROIは、タスク誘発活性化ネットワーク(「TIA」)に結合された。 信号変化が減少した領域上のROIは、タスク誘発性不活性化ネットワーク(「TID」、表1、2)で結合された。 可能な限り、ROI名はAALアトラス(Tzourio-Mazoyerら、2002)を使用して決定した。
表2および図2は、すべてのROIのサイズおよび位置の概要を提供する。 ROIのサイズと境界はあらかじめ定義されているため、例えばROI内の活性化ボクセルの数が比較的少ないために、ROI内の平均活性がベースラインと有意に異ならない可能性がある。 ネットワークレベルの平均活性は、すべての条件でベースラインから有意に異なるが(表3)、CueEでは31個の個別ROIのうち8個、CueDでは31個のROIのうち3個はそうなっていない(表4)。 すべての参加者の信号を含むボクセルのみが解析に含まれた。 ROIの最大サイズは125ボクセルであり、いくつかのROIは頭蓋骨の近くに配置されているため、125ボクセル未満を含む。 関心領域(ROI)位置と全脳fMRI結果の概要。 (A)課題誘発活性化(赤)と課題誘発不活性化(青)のROI 注:ROI22は選択したスライスの外にあるため表示されていない。 MNI座標、BA番号、ROIの大きさは表2にある。 (B)CueE対ベースラインのT値の視覚的表現(≧3=赤→黄、≦-3=青→水色)、(C)CueD対ベースラインのT値(≧3=赤→黄、≦-3=青→水色)。 画像は神経学的方向(L=左、R=右)、名前、略語、ピーク活性化ボクセルのMNI座標は、表1に記載されている。 ネットワーク結果。
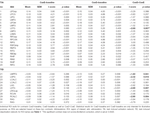
Table 4. ROIの結果。
Hypotheses Testing
本研究では以下の仮説を検証した。
仮説1:TIAはCueEと比較して、CueDの信号変化が増加する。
仮説2:TIDはCueEと比較してCueDの信号変化が減少する。
統計解析
この特定の課題デザインが参加者に初めて提示されたため、パワー分析は行わなかった。 したがって、予想される信号変化率や条件のばらつきに関する知識はない(Desmond and Glover, 2002)。
タスク精度は、StimEとStimDについて正しい反応の割合として計算された。 反応時間は、StimEとStimDについて、全正解数で計算した。
CueEとCueDのベースラインと比較したTIAとTIDの信号変化率の比較には1標本のt検定を使用した。 CueDとCueEの信号変化、精度、反応時間の差の比較には、対の標本片側t検定を用いた(仮説1、仮説2)。 ネットワークオムニバステストが有意でない場合、ROI解析はボンフェローニ補正を行った。 補正前のp値と、このp値がすべての多重比較でボンフェローニ補正に耐えたかどうかを示すコメントを表示する。
Results
Performance
簡単な刺激(「StimE」)の精度は98% ± 1.2% SEM、難しい刺激(「StimD」)は85% ± 2.6%であった。 反応時間は、StimEが682±20ms、StimDが1,138±27msであった。 両条件とも、タスクが参加者全員の指示通りに実行されたと確信できる十分な正確率を示した。 仮説通り、参加者はStimEよりもStimDの方が精度(T = -4.3, p < 0.001)と反応が遅く(T = 17.8, p < 0.001)、易しい刺激と難しい刺激の対比が成功したことがわかる(図3)
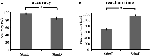
Figure 3. パフォーマンス結果。 (A)両刺激カテゴリーの精度、(B)両刺激カテゴリーの反応時間。 エラーバーは平均の標準誤差(SEM)を表す。 *p ≤ 0.001. 略語。 略号:StimE:簡単な刺激を示す手がかりの後に簡単な刺激を与える、StimD:難しい刺激を示す手がかりの後に難しい刺激を与える。
Functional MRI
Descriptives
TIAネットワークは20のROIからなり、そのうち10はCueEの局所最大値、5はCueDの局所最大値、5はCueEとCueD両方の最大値を含んでいた(表1)。 これら20のROIのうち、CueEでは19、CueDでは18がベースラインと比較して有意な活性を示した(表1)。 TIAにおけるベースラインからの信号変化の平均値は、CueEで0.12 ± 0.02 (t = 6.11, p < 0.001) 、CueDで0.12 ± 0.02 (t = 7.61, p < 0.001) であった。
TIDネットワークは11のROIからなり、そのうち3つはCueEの局所最大値に基づいており、6つはCueDの局所最大値に基づいており、2つはCueEとCueDの両方の最大値を含んでいた(表1)。 これらの11のROIのうち、CueEでは4つ、CueDでは10つがベースラインと比較して有意な活動を示した(表1)。 TIDにおけるベースラインからの信号変化の平均値は、CueEで-0.07 ± 0.02 (t = 2.68, p < 0.01), CueDで -0.12 ± 0.02 (t = 5.37, p < 0.001; 表3)であった。
期待される難易度への同調
TIAの信号増加はCueDではCueEに比べて有意に強くなく(t = -0.24, p = 0.41)、TIA活性化レベルは期待される難易度に同調しないことが示された(図4、表3)。 ポストホックROI分析では、ボンフェローニ補正を適用した後、CueDがCueEに比べて有意に信号が増加したTIA内の20のROIはなかった(表4、図5)
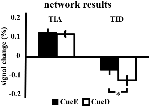
Figure 4. ネットワーク結果の概要。 エラーバーはSEMを表す。 *p ≤ 0.05で、キューが難しい対キューが簡単。 略号は 略号:TIA, task induced activation network; TID, task induced deactivation network; CueE, easy刺激を示す手がかり、刺激が続かない; CueD, difficult刺激を示す手がかり、刺激が続かない
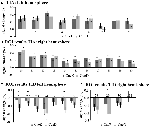
FIGURE 5. 課題誘発活性化ネットワーク内の個々のROI反応の概要、左(A)、右半球(B)、課題誘発不活性化、左(C)、右半球(D)。 エラーバーはSEMを表す。 *CueD vs CueEではp≦0.05、ROIの位置は図2参照、ROIの名称、略称、MNI座標は表1参照。
TIDの信号減少はCueDではCueEに比べて有意に強く(T = -2.48, p = 0.01)予想困難度に調整されることが示された(表3、図4)。 ポストホックROI分析では、TIDの11ROIのうち、CueDではCueEに比べて有意に信号が減少したLMPFG(T = -1.83, p = 0.04), LSPG(T = -2.32, p = 0.02), LSTG(T = -2.69, p < 0.01), RCALC(T = -2.90, p < 0.01)の4つが示されました。 これらの結果は、予想される難易度に対する同調が左前頭、側頭、視覚ROIで最も強いことを示している(表4、図5)
Discussion
本研究では、間歇的覚醒がこれから起こる認知チャレンジの予想難易度に同調するか、どのように同調するかを調べた。 その結果、覚醒度は活性化した脳領域と不活性化した脳領域に反映された。 活性化した領域には、中枢実行ネットワークの領域が含まれた。 また、活性化されたネットワークには、視覚野と同様にDMNの領域が含まれていた。 また、脳の不活性化の度合いは、これから受ける刺激の難易度に比例していたが、活性化の度合いは比例していなかった。 これらの結果は、位相的覚醒の調節は、現在の課題とは無関係な段階的抑制過程(例えば、心・迷走神経、聴覚、視覚過程)に主に反映されることを示唆する。 3545>位相的覚醒に関するこれまでの画像研究では、難易度を1つにしたり、活性化した脳領域のみを提示したり(Shulmanら、1999;Sturm and Willmes、2001;Thielら、2004;Fanら、2005;Périnら、2010;谷中ら、2010)、視覚野の活動のみに注目したり(Bartolucci and Smith、2011)している。 これらの研究から、手がかりによって活性化されるのは、ほとんどが右側にある前頭葉-頭頂葉-視床のネットワークであり、運動野と視覚野が組み合わされることもあることが明らかになりました。 また、活性化されるネットワークは研究によって大きく異なる。 先行研究のネットワーク間の差のいくつかは、キューの後に行われたタスクで説明できるかもしれない。 例えば、Thielら(2004)は視覚知覚課題を用いて視覚皮質領域の活性化のみを同定し、Fanら(2005)とYanakaら(2010)は運動反応課題を用いて視床および運動領域の活性化を同定しています。 Bartolucci and Smith (2011)は、4段階の難易度を持つエレガントな手がかり課題デザインを提示したが、視覚野の活性化のみを提示している。 したがって、この課題において他の脳領域がどのように反応したかは不明である。 本研究で同定された課題誘発性活性化ネットワークは、同様のタイプの視覚処理課題を用いたShulmanら(1999)が発見したネットワークと最も類似している。
位相性覚醒が非情報性手がかりに対するボトムアップ反応かトップダウン反応かは不明(Thielら、2004;Hackley、2009;Périnら、2010;Bartolucci and Smith、2011;Chicaら、2016)であった。 位相性覚醒は、ボトムアップ型(Sturm et al., 1999; Hackley, 2009)、トップダウン型(Bartolucci and Smith, 2011)、複合型(Thiel et al., 2004; Périn et al., 2010)反応として考えられてきた。 我々は、被験者が外部手がかりとその手がかりに関連する指示の両方に反応することから、手がかりに対する脳の反応をボトムアップとトップダウンの処理の組み合わせとして表示することが最も適切であると主張する。 しかし、我々の主な結果は、異なるタイプの課題刺激を示す2つの類似した手がかりに対する反応の差に関するものである。 したがって、困難な課題を示すキューと容易な課題を示すキューとの間の結果の違いは、キューと関連する指示の解釈に関連するトップダウン制御のレベルを表す可能性があると主張する。
課題誘発不活性化ネットワークにはDMNの脳領域、すなわち内側前頭回、上側頭回、角回(Shulmanら、1999; Raichleら、2001; Raichle、2015)が含まれていた。 DMNの不活性化は、認知的挑戦を妨害する内部プロセスを減らすことによって、課題遂行を助けるという仮説が一般的である(Raichleら、2001;Bucknerら、2008;Raichle、2015)。 この見解は、タスク実行中にDMNの不活性化のレベルがタスクの難易度に比例することを示す研究によって支持される(McKiernanら、2003;SinghおよびFawcett、2008)。 これらの研究は、課題実行中の不活性化を調べたものである。 我々の結果は、このスケーリングがタスク予期段階ですでに起こりうることを示唆している。 Weissmanら(2006)は、個人の反応時間のばらつきを調べ、注意の遅れを特定した。 注意の欠如は、DMNの不活性化の減少と関連している可能性がある。 Weissmanら(2006)は注意レベルの自然な変動に注目したが、我々の結果は、いくつかのDMN領域の不活性化も課題刺激提示前にトップダウン的に調整できることを示した。 以下では、これらの領域の提案された機能との関連で、これらの効果の可能な意味を議論する。
内側前頭前野の活動は、以前に心の迷い(Bertossiら、2017)、記憶の検索(Eustonら、2012)、心肺制御(Wongら、2007)に関連していた。 この内側前頭葉の不活性化は,認知的挑戦を予期してこれらの機能のいずれかが既に抑制されていることを示唆している。
視覚領域と時間領域で予想される困難に対する不活性化の同調は,無関係な視覚や聴覚の入力の抑制に関係している可能性がある。 また、両側の側頭領域における同調は、特にMRIスキャナから発生する騒音の抑制に関連している可能性がある。 音のレベルは両条件で同様であると予想するのが妥当である。 また、難易度の高い手がかりに対して非活性化のレベルが強いということは、困難な課題が予想される場合には、周囲の音の処理がより強く抑制されることを示唆している。 視覚野の脱活性化は、側頭葉の脱活性化に比べてあまり報告されていない。 しかし、Smithら(2000)は、視覚野の広範囲の失活と絵の特定の部分への注意の関連を報告したことがある。 また、Giesbrechtら(2006)は、視野の中心付近にある物体へのひそかな注意と、周辺視野にある物体への注意に関連した視覚活性化の変調を報告しています。 これらの知見は、視覚野の活性化が、中心領域と周辺領域のいずれに注意を向けることによっても調節されることを示しています。 さらに、視覚野の活性化は、視覚的注意の位置だけでなく、視覚的チャレンジの予想難易度によっても調節されることを示唆し、また、手がかりの位置とサイズが同じであることから、このプロセスはトップダウン方式で制御されていると思われた。 このネットワークは、以前にワーキングメモリ課題の実行に関連した中央実行ネットワークと類似性を示した(Lawrence et al., 2003)。 しかし、このネットワークの活性化は、予想される課題の難易度に同調するものではありませんでした。 このことは、これらの領域が、困難な条件と容易な条件とで類似のプロセスを表していることを示唆している。 このネットワークは、手がかりの評価、あるいは課題の文脈や指示の検索と維持に関連している可能性がある。 まず、手がかりが先行しない刺激を含む試行を含まなかったため、認知パフォーマンスに対する手がかりの効果を検討することはできない。 また、時間的な制約から、同じ刺激で手がかりのない課題を繰り返すことはできなかった。
次に、困難な認知的課題を示す手がかりの一部として感嘆符を用いたが、この記号は通常、注意力を高める必要がある場合に使用されるものである。 感嘆符は、その固有の意味から、簡単な条件と難しい条件とで異なる活性化パターンに関与している可能性がある。 しかし、このことは、我々の主な結果がトップダウン反応ではなくボトムアップ反応に分類される可能性があることを除けば、我々の結果の主な解釈には影響しない。 最後に、我々の実験デザインは、視覚・知覚的認知課題のみを含むため、位相的覚醒に関連する活動が他の領域に一般化できるか、あるいは領域特異的であるかを検証することはできない<3657><3545>結論として、本研究では、位相的覚醒の同調はいくつかの領域における不活性化のレベルにより表されることを示した。 この効果は内側前頭前野、視覚野、側頭葉で最も強く、心肺制御の抑制、周辺視野の視覚処理、MRIスキャナのノイズの抑制を反映していると思われる。 これらの結果から、認知パフォーマンスは、これから起こる認知的チャレンジの難しさに対する期待と密接に結びついた脳の状態によって促進されることが示唆された。
Data Availability
生データはElisabeth-Tweesteden病院で作成されました。 本研究の知見を裏付ける派生データは、妥当な要求があれば対応する著者から入手できる。
Ethics Statement
この研究は、オランダ社会支援法、医療倫理委員会ブラバントの勧告に従って実施された。 すべての被験者はヘルシンキ宣言に基づき、書面でのインフォームドコンセントを行った。 プロトコルは医療倫理委員会により承認された。 MDとISが実験を行った。 G-JR、NR、JJが共同監修した。 また、G-JR、NR、JJが共同監修し、全著者が査読を行った。
Conflict of Interest Statement
著者らは、本研究が潜在的な利益相反と解釈される商業的または金銭的関係がない状態で行われたことを宣言する。
Acknowledgements
この研究に貢献した放射線技師William PigmansとMaikel Brandsに感謝したい。
略語
DMN, default mode network; M, male; F, female; GLM, general linear model; SPM, statistical parametric mapping; CueE, easy stimulus, not followed by stimulus; CueD, difficult stimulus, not followed by stimulus.易しい刺激を示す手がかりを示す手がかりで、その後に刺激が続かない。 StimE:簡単な刺激の後に簡単な刺激が続く、StimD:難しい刺激の後に難しい刺激が続く、MRI:磁気共鳴イメージング、ROI:関心領域、GDL:GNUデータ言語、TIA:課題誘発活性化ネットワーク、TID:課題誘発非活性化ネットワーク、SEM:平均の標準誤差。
Footnotes
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M…, およびSmith, A. T. (2011). 視覚野における注意の変調は知覚学習中に修正される。 Neuropsychologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi、 E., Peccenini、 L., Solmi, A., Avenanti, A. and Ciaramelli、 E. (2017)(2017). 内側前頭前野の経頭蓋直流刺激は、男性の心の迷いを減衰させる。 Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., and Schacter, D. L. (2008). 脳のデフォルトネットワーク:解剖学、機能、および疾患との関連性。 Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Buracas, G. T., and Boynton, G. M. (2002). M系列を用いた事象関連fMRI実験の効率的な設計。 Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | CrossRef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P., and Bushnell, M. C. (2015). ワーキングメモリータスクの成功には、応答性の高いデフォルトモードネットワークが必要なのか? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | Cross Ref Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., and Paz-Alonso, P. M. (2016). 前頭葉-線条体ネットワークにおける位相的覚醒と意識の相互作用. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | CrossRef Full Text | Google Scholar
Desmond, J. E., and Glover, G. H. (2002). 機能的MRI(fMRI)ニューロイメージング研究におけるサンプルサイズの推定:統計的パワー分析。 J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., and McNaughton, B. L. (2012). 記憶と意思決定における内側前頭前野の役割。 Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract |Ref Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J.I. and Posner, M. I. (2005). 注意ネットワークの活性化。 Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T. , Raz, A. and Posner, M. I. (2002)(FAN, J. , McKandliss, B. D. , Raz, A. , M. I. 注意ネットワークの効率と独立性のテスト。 J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/089892902317361886
PubMed Abstract | CrossRef Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G. and Mangun, G.R. (2006)(2011). 視覚野のターゲット前活動は空間および特徴的注意課題における行動成績を予測する。 Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009).視覚野におけるターゲット前の活動は、空間的および特徴的な注意のタスクの行動パフォーマンスを予見する。 警告信号による自発的反応の速まり。 Psychophysiology 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R. , Sorsa, M., and Siivonen, P. (2008). 橋渡し役における6/6および4/8当直制の眠気に対する効果。 Chronobiol. 25, 413-423. doi: 10.1080/07420520802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A., and Buckner, R. L. (2012). 白質病理と関連した進行した老化における注意力制御の失敗。 Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., and Duyn, J. H. (2007). fMRI study of effort and information processing in a working memory task.(作業記憶タスクの作業と情報処理に関するfMRI研究). Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., and Rutten,G. J. (2017). “A proposal for a system that facilitates quantitative replication of fMRI studies,” in Proceedings of the Human Brain Mapping Conference 2017 June 25-29, Vancouver, Canada.「fMRI研究の定量的な再現を促進するシステムの提案」. オンラインで入手可能。 https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., and Baker, C. I. (2009).を参照。 システム神経科学における円形解析:二重浸漬の危険性。 Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A., and Bartolomeo, P. (2011). 位相的な聴覚的注意喚起は視覚的意識知覚を改善する。 Conscious. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G., and Stein, E. S. (2003)(2013). 複数の神経ネットワークが持続的注意を媒介する。 J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/089892903770007416
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, G., Sobering, G., Duyn, J. and Moonen, C. T. (1993). エコーシフトの原理と観察列を組み合わせた機能的MRI技術(PRESTO)。 Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | CrossRef Full Text | Google Scholar
Macleod, J.W., Lawrence, M.A., McConnell, M.M., Eskes, G.A., Klein, R.M., and Shore, D.I. (2010). ANTの評価:注意ネットワークテストの心理測定と理論的考察。 Neuropsychology 24, 637-651. doi: 10.1037/a0019803
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K.A., D’Angelo, B.R.D., Kaufman, J.N. and Binder, J.R. (2006). 意識の流れ」に割り込む:fMRIによる検討。 Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K.A., Kaufman, J.N., Kucera-Thompson, J.K., and Binder, J. R.(2003)。 機能的ニューロイメージングにおける課題誘発性不活性化に影響を与える要因のパラメトリック操作。 J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/0898929033593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R.D., and Brady, C.B. (1993). アルツハイマー病におけるphasic and tonic alertness。 Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., and Ramsey, N. F. (2008)(Needs, R. F. (2008). 高速3次元血液酸素レベル依存型機能的MRIによる感度の向上:3 TでのSENSE-PRESTOと2D-EPIの比較 NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., and de Marco, G. (2010). 健康な若年被験者における注意力:警告信号によって増強される脳領域の相互作用のfMRI研究。 Brain Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H.他 (2009).Al. ワーキングメモリ負荷がその後の安静相におけるFMRI安静状態パターンに与える影響。 PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract |Ref Full Text | Google Scholar
Raichle, M. E. (2015). 脳のデフォルトモードネットワーク. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | Cross Full Text | Google Scholar
Raichle, M.E., MacLeod, J.W., Snyder, A. Z., Powers, W.J., Gusnard, D.A., and Shulman, G. L. (2001). 脳機能のデフォルトモード。 Proc. Natl.Acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M. , Akbudak, E., Conturo, T. E. , Snyder, A. Z., Petersen, S. E. , et al. (1999). 移動物体に対する方向期待の符号化と適用に関与する領域。 J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K.D., and Fawcett, I.P. (2008). 視覚的検出課題によるヒトのデフォルトモードネットワークの一過性および線形的な勾配性不活性化。 Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith,A. T., Singh, K. D., and Greenlee, M. W. (2000). ヒト視覚野における注意の活動抑制。 Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W.de Simone, A., Krause, B. J., Specht, K.Hessemann, V., Radermacher, I., et al.(1999).のような、視覚野における注意力抑制の研究。 内的覚醒の機能解剖学:右半球のフロート-頭頂-視床-脳幹ネットワークに関する証拠。 Neuropsychologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., and Willmes, K.(2001). 内在的覚醒と位相的覚醒の機能的神経解剖学について。 Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | CrossRef Full Text | Google Scholar
Thiel, C. M., Zilles, K., and Fink, G. R. (2004)(2004. 視空間的注意の覚醒、方向づけ、再方向づけの大脳相関:事象関連fMRI研究. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O. , Delcroix, N., et al. MNI MRI単一被験者脳の巨視的解剖学的区画を用いたSPMにおける活性化の自動的解剖学的ラベリング。 Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | CrossRef Full Text | Google Scholar
van Gelderen, P., Ramsey, N.F., Liu, G., Duyn, J.H., Frank, J.A., Weinberger, D. R.、他 (1995).The Maskical Parcellation of the MNI MRI Single Subject Brain. 臨床用1.5Tスキャナーによるヒト脳の3次元機能的磁気共鳴画像。 Proc. Natl.Acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N. and Henik, A. (2011). 位相的覚醒は、視覚刺激のグローバルな処理を強化することにより、実行制御を調節することができる。 Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., Visscher, K. M., and Woldorff, M. G. (2006). 注意の瞬間的な欠落の神経基盤。 Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | CrossRef Full Text | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S., and Shoemaker, J. K. (2007)(2007. 意識下のヒトにおける腹内側前頭前野と心拍数制御。 Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka,H. T., Saito,D. N., Uchiyama,Y. and Sadato, N. (2010). 位相性覚醒の神経基盤:機能的磁気共鳴画像法による研究. Neurosci. 斉藤大輔・内山靖弘・貞藤憲史(2010):位相性覚醒の神経基盤:機能的磁気共鳴画像法による検討.