Introduction
Aiemmat tutkimukset osoittavat, että ihmiset voivat käyttää vihjeitä valmistautuakseen henkisesti kognitiiviseen haasteeseen. Jos osallistujia ohjeistetaan valmistautumaan henkisesti sillä hetkellä, kun vihje esitetään, tämä on määritelty vaiheelliseksi valppaudeksi . Kyky vaikuttaa vireystasoon voi olla tärkeä, erityisesti jos vireystasolla on myös hintansa. Jos vireystilan vaatimus on liian suuri, se voi mahdollisesti johtaa väsymykseen (Härmä ym., 2008) tai stressiin liittyviin ongelmiin. Tässä tutkimuksessa pyrimme selvittämään, virittävätkö terveet aikuiset vireystasonsa tulevan kognitiivisen tehtävän odotettuun vaikeuteen ja miten he sen virittävät.
Jos vihje esitetään juuri ennen tehtävän ärsykettä, ihmiset pystyvät yleensä reagoimaan nopeammin yksinkertaisissa motorisissa vastaustehtävissä (Fan ym., 2002; Macleod ym., 2010; Weinbach ja Henik, 2011) ja hahmottamaan heikentyneet visuaaliset ärsykkeet tarkemmin (Kusnir ym., 2011). Kuvantamistutkimukset ovat osoittaneet, että vaiherikas vireystila liittyy lisääntyneeseen aktivaatioon otsa-, parietaali-, talamuksen (Shulman ym., 1999; Fan ym., 2005; Yanaka ym., 2010), temporo-okzipitaali- (Thiel ym., 2004) ja visuaalisilla aivoalueilla (Bartolucci ja Smith, 2011). Nämä kuvantamistutkimukset ovat antaneet arvokasta tietoa vaiheittaisen vireystilan edustuksesta aivoissa. Faasista vireystilaa on kuitenkin yleensä tutkittu on/off-ilmiönä tai analyysi on rajoitettu näköaivokuoreen. Siksi on epäselvää, heijastaako vireystila myös kognitiivisen haasteen odotettua vaikeutta koko aivoverkostoissa.
Lisäksi aiemmat tutkimukset ovat osoittaneet, että oletustilaverkostoon (default mode network, DMN) kuuluvien aivoalueiden deaktivointi voi helpottaa kognitiivisen tehtävän suorittamista. Ensinnäkin tehtävien suorittamista koskevat tutkimukset osoittavat, että kasvava tehtävän vaikeustaso liittyy lisääntyvään deaktivoitumiseen (McKiernan et ai., 2003, 2006; Jansma et ai., 2007; Singh ja Fawcett, 2008; Pyka et ai., 2009; Hedden et ai., 2012; Čeko et ai., 2015). Toiseksi Jansma et al. (2007) ovat osoittaneet, että DMN:n mediaalinen prefrontaalinen osa virittyy vain tehtävän vaikeuteen, jonka osallistuja pystyi ennakoimaan, mutta siihen ei vaikuttanut kunkin ärsykkeen todellinen vaikeus, jota osallistuja ei voinut ennakoida. Lopuksi Weissman et al. (2006) havaitsivat, että DMN:n deaktivoituminen juuri ennen ärsykettä oli heikompaa, jos koehenkilöt vastasivat suhteellisen hitaasti. Ehkä nämä koehenkilöt vastasivat hitaammin, koska he olivat tilapäisesti vähemmän valppaita.
Tutkimustamme varten suunnittelimme cued visuaalisen havaintotehtävän, jossa oli helppo ja vaikea ehto. Vihjeet antoivat tietoa tulevan ärsykkeen vaikeudesta, mutta eivät antaneet mitään tietoa, joka helpottaisi itse tehtävää. Puolessa kokeista vihjettä ei seurannut ärsyke. Ainoastaan nämä kokeet analysoitiin aivotoiminnan osalta, jotta voitiin täysin eristää vihjeeseen liittyvä aktivaatio tehtävän suorittamiseen liittyvästä aktivaatiosta. Oletamme, että osallistujat virittävät aktiivisuutensa tehtävän odotetun vaikeuden mukaan.
Materiaalit ja menetelmät
Osallistujat
Osallistujat rekrytoitiin verkkomainonnalla. Osallistujat suljettiin pois, jos heillä oli aiemmin ollut merkittäviä neurologisia tai psykiatrisia häiriöitä tai jos heillä oli vasta-aiheita magneettikuvaukselle (metalliesineet kehossa tai sen ympärillä, klaustrofobia tai raskaus). Tämä tutkimus toteutettiin ihmisiin kohdistuvaa lääketieteellistä tutkimusta koskevan lain (WMO), Brabantin lääketieteellisen tutkimuksen eettisen komitean suositusten mukaisesti. Lääketieteellisen tutkimuksen eettinen komitea hyväksyi tutkimussuunnitelman (protokollanumero: NL51147.028.14). Kaikki koehenkilöt antoivat kirjallisen tietoon perustuvan suostumuksen Helsingin julistuksen mukaisesti.
Tutkimukseen osallistui 22 tervettä oikeakätistä vapaaehtoista. Kahden osallistujan tiedot suljettiin pois skannerin artefaktien vuoksi. Tulokset raportoidaan jäljelle jääneistä 20 osallistujasta.
Tehtävän suunnittelu
Suunnittelimme tehtävän (tapahtumakohtaisella suunnittelulla), jonka avulla pystyimme tutkimaan tehtävän odotettuun vaikeuteen liittyviä ennakointivaikutuksia ilman tehtävän suorittamisen sekoittavia vaikutuksia.
Tehtävän ärsykkeet koostuivat yhdeksästä nuolesta, jotka olivat kolmella kolme kertaa kolmella sijoittelevassa asettelussa mustalla kuvaruudulla (kuva 1). ”Helppo ärsyke” sisälsi kahdeksan nuolta oikeaan suuntaan ja yhden nuolen vastakkaiseen suuntaan. Vaikea ärsyke sisälsi viisi nuolta oikeaan suuntaan ja neljä nuolta vastakkaiseen suuntaan (kuva 1). Osallistujia ohjeistettiin painamaan nuolten enemmistön suuntaa vastaavalla kädellä nappia. Helpot ja vaikeat ärsykkeet esitettiin satunnaisessa järjestyksessä. Perusärsykkeet koostuivat paikallaan olevasta mustasta ruudusta, jossa oli teksti: ”sinulla on 30 sekunnin tauko” hollanniksi ja valkoisin kirjaimin. Ärsykkeet esitettiin ”presentation”-ohjelmistolla.
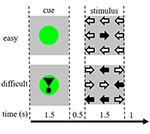
Kuva 1. Visuaalisten ärsykkeiden esitys. Kukin koe kesti 4,5 s. Puolessa kokeista vihjettä ei seurannut ärsyke. Näitä kokeita käytettiin funktionaalisen magneettikuvauksen (fMRI) analyysiin.
Jokainen koe alkoi vihjeellä, joka osoitti myöhemmin esitettävän ärsykkeen vaikeuden. Vihreä piste osoitti ”helppoa ärsykettä”, vihreä piste huutomerkillä osoitti ”vaikeaa ärsykettä” (kuva 1).
Puolessa kokeista vihjettä ei seurannut ärsyke, toisessa puolessa ärsyke esitettiin 500 ms sen jälkeen, kun vihje oli kadonnut. Näin saatiin neljä olosuhdetta: (1) helppoa ärsykettä koskeva vihje, jota ei seurannut ärsyke (”CueE”); (2) vaikeaa ärsykettä koskeva vihje, jota ei seurannut ärsyke (”CueD”); (3) helppoa ärsykettä koskeva vihje, jota seurasi helppo ärsyke (”StimE”); ja (4) vaikeaa ärsykettä koskeva vihje, jota seurasi vaikea ärsyke (”StimD”). Kaikki olosuhteet erotettiin lähtötilanteesta. Toiminnallisen magneettikuvauksen (fMRI) tulokset perustuivat CueE:hen ja CueD:hen. Suorituskykytulokset perustuivat StimE:hen ja StimD:hen. CueE ja CueD esitettiin pseudosattumanvaraisessa järjestyksessä käyttäen M-sekvenssejä yleisen lineaarisen mallin (GLM) regressorin riippumattomuuden maksimoimiseksi olosuhteiden välillä ja suunnittelun tehokkuuden optimoimiseksi (Buracas ja Boynton, 2002). Osallistujat eivät voineet ennustaa, seuraisiko vihjettä ärsyke vai ei.
Kunkin kokeen kesto oli 4500 ms. Vihjeet esitettiin t = 0:ssa 1 500 ms:n ajan ja ärsykkeet esitettiin t = 2 000:ssa, myös 1 500 ms:n ajan (kuva 1). Tehtävä suoritettiin kahdessa 68 kokeen jaksossa. Lähtötilanne koostui kolmesta 30 sekunnin lepojaksosta ennen kahta suoritusta, sen jälkeen ja niiden välissä. Tehtävän kokonaiskesto oli 11,7 minuuttia. Tämä oli istunnon toinen tehtävä, ja se alkoi sen jälkeen, kun osallistujat olivat makailleet noin 10 minuuttia skannerissa.
Osallistujia ohjeistettiin valmistautumaan helppoon tai vaikeaan ärsykkeeseen vihjeen tietojen perusteella (kuva 1) ja osoittamaan mahdollisimman tarkasti ja nopeasti ärsykkeen useimpien nuolien suunta. Osallistujat harjoittelivat tehtävää skannerin ulkopuolella 3 minuutin mittaisen standardiharjoitusprotokollan mukaisesti. Tämä protokolla toistettiin, jos suoritus oli riittämätön (alle 70 % tarkkuus vaikeiden ärsykkeiden kohdalla).
Pääkäämiin kiinnitetyn peilin avulla osallistujat pystyivät näkemään pään taakse sijoitetun läpinäkyvän projektionäytön. Skannerihuoneessa oleva videoprojektori heijasti tehtäväärsykkeet näytölle. Reaktioiden tallentamiseen käytettiin kahta paineilmalla toimivaa pneumaattista painonappilaatikkoa. Osallistujat käyttivät vasenta peukaloa tai etusormea vastatessaan ”vasemmalle” ja oikeaa vastatessaan ”oikealle.”
Kuvan hankinta
Kuvaukset suoritettiin 3T Philips Achieva -skannerissa (Philips Medical Systems, Best, Alankomaat) käyttäen 32-kanavaista SENSE-pääkäämiä. Anatomista rekisteröintiä varten otettiin 3D T1-painotteinen rakennekuva . fMRI-kuvat saatiin käyttämällä 3D PRESTO-pulssisekvenssiä . Skannerista otettiin kuusi tyhjää kuvaa, jotka skanneri hylkäsi.
Kuvan esikäsittely
fMRI-tiedot esikäsiteltiin käyttämällä tilastollista parametrikartoitusta (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, Lontoo, UK2). Yhden istunnon skannaukset sovitettiin uudelleen ensimmäiseen skannaukseen koehenkilön liikkeen korjaamiseksi käyttäen pienimmän neliösumman lähestymistapaa, kuuden parametrin (jäykkä keho) spatiaalista muunnosta ja 2. asteen B-spline-estimointia. Skannaukset rinnakkaisrekisteröitiin T1:een käyttämällä jäykän kehon mallia. Parametrit estimoitiin normalisoidun keskinäisen informaatiofunktion avulla. Kuvat leikattiin uudelleen 4. asteen B-splinellä. T1 normalisoitiin spatiaalisesti tavanomaiseen MNI-avaruuteen käyttäen hyvin kevyttä bias-regularisointia (0,0001) ja 4. asteen B-splinea. Tuloksena saatuja parametreja sovellettiin kaikkiin toiminnallisiin skannauksiin anatomisten erojen minimoimiseksi ja siten ryhmäanalyysin mahdollistamiseksi. Lopuksi kaikki skannaukset tasoitettiin spatiaalisesti 3D-Gaussin suodattimella (täysleveys puolimaksimissa: 8 mm) toiminnallisten anatomisten erojen vaikutuksen minimoimiseksi entisestään.
Yksilöllinen fMRI-analyysi
Yksilöllinen GLM-regressioanalyysi suoritettiin kullekin vokselille yksilöllisten aktivointikarttojen luomiseksi käyttäen massan-yksilomuuttujaa, jossa käytettiin globaalia likimäärin arvioitua AR: n (1) autokorrelaatiomallia ja ylipäästösuodatinta (128 s:n katkaisu). Lähtötasoa ei nimenomaisesti mallinnettu. Perusfunktio oli kanoninen HRF ilman derivaattoja. Peittokynnyksenä käytettiin 0,8. Kullekin tilalle käytettiin erillisiä regressoreita (CueE, 17 aikapistettä; CueD, 16 aikapistettä; StimE, 17 aikapistettä; StimD, 18 aikapistettä; ja häiritsevä regressori m-sekvenssiin liittyvän tehtävän blanc-jaksoille, 68 aikapistettä). Analyysistä ei jätetty pois yhtään aikapistettä. Beeta-kartat muunnettiin vastaamaan todellista prosentuaalista signaalimuutosta kussakin vokselissa. Esitämme vain CueE- ja CueD-regressorien tulokset, jotka heijastavat signaalimuutoksia vihjeiden osalta ilman ärsykettä. Beeta- ja tilastolliset t-kartat tarkistettiin visuaalisesti suurten artefaktien varalta.
Ryhmän fMRI-analyysi
Toisen tason fMRI-analyysi suoritettiin lineaarisella regressiolla jokaisessa voxelissa visualisointia ja kiinnostavan alueen (ROI) valintaa varten. ROI-analyysi suoritettiin GNU-datakielellä (GDL3) käyttäen GLM-analyysin tuottamia yksittäisen kohteen prosentuaalisia signaalimuutoskarttoja. ROI:n tai verkon sisällä olevat voxelit keskiarvoistettiin CueE:n, CueD:n ja lähtötason välisen signaalieron laskemiseksi.
ROI-valinta
Supratentoriaaliset paikalliset maksimit ja minimit määritettiin sekä CueE:lle että CueD:lle SPM:llä korjaamattomalla kynnysarvolla (p < 0.001; Taulukko 1). Ennalta määritellyn kokoiset ja muotoiset kuutiomaiset ROI:t sijoitettiin CueE-peruskontrastin ja CueD-peruskontrastin paikallisten maksimien päälle erikseen ennalta määritellyn rasterin sisällä (Jansma ja Rutten, 2017; Taulukko 2). ROI:t otettiin mukaan analyysiin, jos ne sisälsivät merkittävän paikallisen maksimin CueE- tai CueD-kontrastille. Määrittelemällä ROI:t molempien olosuhteiden paikallisille maksimeille erikseen estämme harhaa joko CueE: n tai CueD: n suuntaan. Lisäksi ROI:iden ennalta määritelty muoto ja koko rasterin sisällä vähentää ympyrämäisyyden vaikutusta, koska kohina ei vaikuta näiden ROI:iden rajoihin (Kriegeskorte et al., 2009). Lisäksi tämä menetelmä helpottaa tulosten kvantitatiivista vertailua olosuhteiden välillä ja mahdollisesti eri tutkimusten välillä, mikä helpottaa fMRI-tulosten kvantitatiivista toistettavuutta. ROI:iden sijoittaminen paikallisten maksimien päälle optimoi tehon. Aktiivisuushuipun sijainnin tiedetään kuitenkin vaikuttavan kohinaan, joten ROI-alueiden tarkka sijainti voi aina toistua kunkin ROI-alueen osalta. Tämä ei vaikuta verkkotuloksiin, koska kaikki ROI:t on keskiarvoistettu ja tarkka sijainti ei näin ollen ole yhtä merkityksellinen.
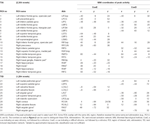
Taulukko 1. Verkkotulokset. Toimintamalli.
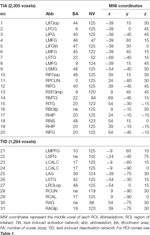
TAULUKKO 2. Kiinnostuksen kohteena olevien alueiden (ROI) ominaisuudet.
ROI:t alueilla, joilla signaalimuutos lisääntyi, yhdistettiin tehtävän aiheuttamaksi aktivaatioverkoksi (”TIA”). ROI-alueet alueilla, joilla signaalimuutos väheni, yhdistettiin tehtävän aiheuttamaksi deaktivointiverkoksi (”TID”; Taulukot 1, 2). ROI-nimet määritettiin mahdollisuuksien mukaan AAL-atlaksen avulla (Tzourio-Mazoyer et al., 2002).
Taulukossa 2 ja kuvassa 2 esitetään yleiskuva kaikkien ROI:iden koosta ja sijainnista. Koska ROI:iden koko ja rajat on määritelty ennalta, on mahdollista, että ROI:n keskimääräinen aktiivisuus ei poikkea merkittävästi lähtötasosta esimerkiksi siksi, että ROI:n sisällä olevien aktivoituneiden vokseleiden määrä on suhteellisen pieni. Vaikka keskimääräinen aktiivisuus verkkotasolla eroaa merkittävästi lähtötasosta kaikissa olosuhteissa (taulukko 3), näin ei ole kahdeksan yksittäistä ROI:ta 31:stä CueE:n osalta ja kolme ROI:ta 31:stä CueD:n osalta (taulukko 4). Analyysiin otettiin mukaan vain ne vokselit, jotka sisälsivät signaalia jokaisen osallistujan osalta. ROI:n maksimikoko on 125 voxelia, jotkin ROI:t sisältävät vähemmän kuin 125 voxelia, koska ne sijaitsevat lähellä kalloa.
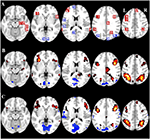
KUVIO 2. ROI:n enimmäiskoko. Yleiskatsaus kiinnostuksen kohteiden (ROI) sijaintiin ja koko aivojen fMRI-tuloksiin. (A) ROI-alueet tehtävän aiheuttamalle aktivaatiolle (punainen) ja tehtävän aiheuttamalle deaktivoitumiselle (sininen) Huomautus: ROI-aluetta 22 ei näytetä, koska sijainti on valittujen viipaleiden ulkopuolella. MNI-koordinaatit, BA-numero ja ROI:iden koko löytyvät taulukosta 2. (B) Visuaalinen esitys t-arvoista CueE:lle vs. lähtötaso (≥3 = punainen keltaiseksi, ≤ -3 = sininen vaaleansiniseksi), (C) T-arvot CueD:lle vs. lähtötaso (≥3 = punainen keltaiseksi, ≤ -3 = sininen vaaleansiniseksi). Kuvat ovat neurologisessa orientaatiossa (L = vasen, R = oikea), huippuaktivaatioiden vokseleiden nimet, lyhenteet, MNI-koordinaatit löytyvät taulukosta 1.

Taulukko 3. Verkkotulokset.
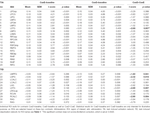
TAULUKKO 4. ROI-tulokset.
Hypoteesien testaus
Testasimme tässä tutkimuksessa seuraavia hypoteeseja:
Hypoteesi 1: TIA osoittaa lisääntynyttä signaalimuutosta CueD:n osalta verrattuna CueE:hen.
Hypoteesi 2: TID osoittaa vähentynyttä signaalimuutosta CueD:n osalta verrattuna CueE:hen.
Statistinen analyysi
Tehoanalyysiä ei suoritettu, koska tämä oli ensimmäinen kerta, kun tämä erityinen tehtäväsuunnitelma esitettiin osallistujille. Siksi ei ole tietoa odotetusta prosentuaalisesta signaalimuutoksesta ja olosuhteiden vaihtelusta (Desmond ja Glover, 2002).
Tehtävän tarkkuus laskettiin StimE:lle ja StimD:lle oikeiden vastausten prosenttiosuutena. Reaktioajat laskettiin StimE:lle ja StimD:lle kaikista oikeista vastauksista.
Yhden otoksen t-testejä käytettiin vertaamaan TIA:n ja TID:n prosentuaalista signaalimuutosta lähtötasoon verrattuna CueE:n ja CueD:n osalta. Parillisten otosten yksipuolisia t-testejä käytettiin vertaamaan signaalimuutosta, tarkkuutta ja reaktioaikojen eroja CueD:n ja CueE:n välillä (hypoteesit 1 ja 2).
Post hoc -testaus sisälsi erilliset t-testit kullekin ROI:lle kussakin verkossa. ROI-analyysi oli Bonferroni-korjattu, jos verkon omnibus-testi ei ollut merkitsevä. Esitämme korjaamattomat p-arvot ja kommentin, jossa todetaan, olisiko tämä p-arvo selvinnyt Bonferroni-korjauksesta kaikkien moninkertaisten vertailujen osalta. Kaikkiin tilastollisiin analyyseihin käytettiin SPSS 24 -ohjelmaa.
Tulokset
Suorituskyky
Tarkkuus helppojen ärsykkeiden (”StimE”) osalta oli 98 % ± 1,2 % SEM ja vaikeiden ärsykkeiden (”StimD”) osalta 85 % ± 2,6 %. Reaktioaika StimE:lle oli 682 ± 20 ms ja StimD:lle 1 138 ± 27 ms. Molempien olosuhteiden tarkkuusasteet ovat riittävät, jotta voidaan olla varmoja siitä, että kaikki osallistujat suorittivat tehtävän ohjeiden mukaisesti. Kuten oletettiin, osallistujat vastasivat epätarkemmin (T = -4.3, p < 0.001) ja hitaammin (T = 17.8, p < 0.001) StimD:ssä kuin StimE:ssä (kuva 3), mikä osoittaa, että kontrasti helppojen ja vaikeiden ärsykkeiden välillä oli onnistunut.
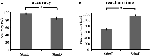
Kuva 3. Suorituskyvyn tulokset. (A) Tarkkuus molemmille ärsykeluokille, (B) reaktioaika molemmille ärsykeluokille. Virhepalkit edustavat keskiarvon keskivirhettä (SEM). *p ≤ 0.001. Lyhenteet: StimE, helppoa ärsykettä osoittava vihje, jota seuraa helppo ärsyke; StimD, vaikeaa ärsykettä osoittava vihje, jota seuraa vaikea ärsyke.
Funktionaalinen magneettikuvaus
Descriptives
TIA-verkko koostui 20 ROI:sta, joista 10 perustui CueE:n paikallisiin maksimiin, viisi perustui CueD:n paikallisiin maksimiin ja viisi sisälsi sekä CueE:n että CueD:n maksimin (taulukko 1). Näistä 20 ROI-alueesta 19:ssä oli merkittävää aktiivisuutta lähtötasoon verrattuna CueE:n osalta ja 18:ssa CueD:n osalta (taulukko 1). Keskimääräinen signaalimuutos TIA:ssa verrattuna lähtötasoon CueE:lle oli 0,12 ± 0,02 (t = 6,11, p < 0,001) ja CueD:lle 0,12 ± 0,02 (t = 7,61, p < 0,001).
TID-verkko koostui 11 ROI: sta, joista kolme perustui CueE: n paikallisiin maksimiin, kuusi perustui CueD: n paikallisiin maksimiin ja kaksi sisälsi sekä CueE: n että CueD: n maksimin (taulukko 1). Näistä 11:stä ROI-alueesta neljällä oli merkittävää aktiivisuutta lähtötasoon verrattuna CueE:n osalta ja 10:llä CueD:n osalta (taulukko 1). Keskimääräinen signaalin muutos TID: ssä verrattuna lähtötasoon CueE: lle oli -0.07 ± 0.02 (t = 2.68, p < 0.01) ja CueD: lle -0.12 ± 0.02 (t = 5.37, p < 0.001; Taulukko 3).
Virittyminen odotettuun vaikeuteen
TIA-signaalin nousu ei ollut merkittävästi voimakkaampi CueD: lle verrattuna CueE: hen (t = -0.24, p = 0.41), mikä osoittaa, että TIA-aktivoitumisen taso ei ole viritetty odotettuun vaikeuteen (Kuva 4, Taulukko 3). Post hoc ROI-analyysi osoitti, että yhdelläkään TIA:n sisällä olevista 20 ROI:sta ei ollut merkittävää signaalin nousua CueD:lle verrattuna CueE:hen Bonferroni-korjauksen soveltamisen jälkeen (taulukko 4, kuva 5).
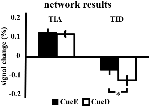
Kuva 4. Yleiskatsaus verkon tuloksiin. Virhepalkit edustavat SEM. *p ≤ 0,05 cue difficult vs. cue easy. Lyhenteet: TIA, tehtävän aiheuttama aktivoitumisverkko; TID, tehtävän aiheuttama deaktivoitumisverkko; CueE, helppoa ärsykettä osoittava vihje, jota ei seuraa ärsyke; CueD, vaikeaa ärsykettä osoittava vihje, jota ei seuraa ärsyke.
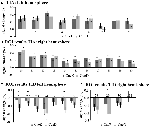
KUVIO 5. Tehtävän aiheuttama aktivoituminen. Yleiskatsaus yksittäisistä ROI-vasteista tehtävän aiheuttamassa aktivoitumisverkossa, vasen (A) ja oikea aivopuolisko (B), ja tehtävän aiheuttama deaktivoituminen, vasen (C) ja oikea aivopuolisko (D). Virhepalkit edustavat SEM:ää. *p ≤ 0.05 CueD vs. CueE, katso kuva 2 ROI: iden sijainnista, katso taulukko 1 ROI: iden nimet, lyhenteet ja MNI-koordinaatit.
Signaalin lasku TID: ssä oli merkittävästi voimakkaampi CueD: lle verrattuna CueE: hen (T = -2.48, p = 0.01), mikä osoittaa, että TID: n taso on viritetty odotettuun vaikeusasteeseen (taulukko 3, kuva 4). Post hoc ROI-analyysi osoitti, että neljällä TID:n 11 ROI-alueesta oli merkitsevä signaalin lasku CueD: lle verrattuna CueE: hen, nimittäin LMPFG (T = -1.83, p = 0.04), LSPG (T = -2.32, p = 0.02), LSTG (T = -2.69, p < 0.01) ja RCALC (T = -2.90, p < 0.01). Nämä tulokset osoittavat, että virittyminen odotettuun vaikeuteen on voimakkainta vasemmanpuoleisissa frontaalisissa, temporaalisissa ja visuaalisissa ROI:issa (Taulukko 4, Kuva 5).
Keskustelu
Tässä tutkimuksessa tutkittiin, viritetäänkö ja miten faasinen valppaus virittyy tulevan kognitiivisen haasteen odotettuun vaikeuteen. Faasinen vireystila heijastui sekä aktivoituneisiin että deaktivoituneisiin aivoalueisiin. Aktivoituneeseen verkkoon kuului keskeisen toimeenpanoverkon alueita. Deaktivoituun verkkoon kuului DMN:n alueita sekä visuaalisia aivokuoria. Deaktivoitumisen taso skaalautui tulevan ärsykkeen odotetun vaikeuden mukaan, kun taas aktivoitumisen taso ei skaalautunut. Nämä tulokset viittaavat siihen, että faasisen vireystilan modulaatio heijastuu pääasiassa asteittaisiin suppressioprosesseihin, joilla ei ole merkitystä nykyisen tehtävän kannalta (esimerkiksi kardio-vagaaliset, auditiiviset ja visuaaliset prosessit). Mahdollisesti tämä tapahtui siksi, että nämä prosessit voisivat häiritä tulevia kognitiivisia haasteita.
Varhaisvaiheen vireystilaa koskevissa aiemmissa kuvantamistutkimuksissa käytettiin yhtä vaikeustasoa, esitettiin vain aktivoituneita aivoalueita (Shulman ym., 1999; Sturm ja Willmes, 2001; Thiel ym., 2004; Fan ym., 2005; Périn ym., 2010; Yanaka ym., 2010) tai keskityttiin vain visuaalisen aivokuoren toimintaan (Bartolucci ja Smith, 2011). Näissä tutkimuksissa on tunnistettu enimmäkseen oikealle lateralisoitunut fronto-parietal-talamuksen verkosto, joskus yhdistettynä motorisiin ja visuaalisiin kortikaalialueisiin, joka aktivoitui vihjeen vaikutuksesta. Tutkimusten välillä on huomattavia eroja aktivoituneissa verkoissa. Osa aiempien tutkimusten verkostojen välisistä eroista saattaa selittyä vihjettä seuranneella tehtävällä. Esimerkiksi Thiel et al. (2004) havaitsivat aktivoitumista vain visuaalisilla kortikaalialueilla visuaalisen havaintotehtävän avulla, kun taas Fan et al. (2005) ja Yanaka et al. (2010) havaitsivat talamuksen ja motoriikan aktivoitumista motorisen vastetehtävän avulla. Vaikka Bartolucci ja Smith (2011) esittävät tyylikkään cued-tehtäväsuunnitelman, jossa on neljä vaikeustasoa orientaatiodiskriminointitehtävässä, he esittivät aktivoitumista vain näköaivokuorella. Siksi on epäselvää, miten muut aivoalueet vastasivat tässä tehtävässä. Tehtävän aiheuttama aktivaatioverkko, joka tunnistettiin tässä tutkimuksessa, muistuttaa eniten Shulmanin et al. (1999) löytämää verkkoa, jossa käytettiin samantyyppistä visuaalista prosessointitehtävää.
On epäselvää, onko vaiheellinen valppaus alhaalta ylöspäin vai ylhäältä alaspäin suuntautuva reaktio epäinformatiiviseen vihjeeseen (Thiel et al., 2004; Hackley, 2009; Périn et al., 2010; Bartolucci ja Smith, 2011; Chica et al., 2016). Faasista valppautta on pidetty joidenkin mielestä alhaalta ylöspäin suuntautuvana (Sturm ym., 1999; Hackley, 2009), toisten mielestä ylhäältä alaspäin suuntautuvana (Bartolucci ja Smith, 2011) ja toisten mielestä yhdistelmävasteena (Thiel ym., 2004; Périn ym., 2010). Väitämme, että on sopivinta nimittää aivovaste vihjeeseen alhaalta ylöspäin ja ylhäältä alaspäin tapahtuvan prosessoinnin yhdistelmäksi, koska osallistujat reagoivat sekä ulkoiseen vihjeeseen että tähän vihjeeseen liittyvään ohjeeseen. Tärkeimmät tuloksemme koskevat kuitenkin eroa vasteessa kahteen samanlaiseen vihjeeseen, jotka osoittavat erityyppistä tehtäväärsykettä. Siksi väitämme, että tuloksena oleva ero vaikeaa tehtävää osoittavan vihjeen ja helppoa tehtävää osoittavan vihjeen välillä voi edustaa ylhäältä alaspäin suuntautuvan kontrollin tasoa, joka liittyy vihjeen ja siihen liittyvän ohjeen tulkintaan.
Tehtävän aiheuttama deaktivointiverkko sisälsi DMN:n aivoalueita, nimittäin mediaalisen prefrontaalisen gyruksen, ylemmän temporaalisen gyrin ja kulmahaarakkeen gyriin (Shulman et ai., 1999; Raichle et ai., 2001; Raichle, 2015). Yleisesti oletetaan, että DMN-deaktivaatio auttaa tehtävien suorittamista vähentämällä sisäisiä prosesseja, jotka häiritsevät kognitiivisia haasteita (Raichle et ai., 2001; Buckner et ai., 2008; Raichle, 2015). Tätä näkemystä tukevat tutkimukset, jotka osoittavat, että DMN-deaktivaation taso skaalautuu tehtävän vaikeuden kanssa tehtävän suorittamisen aikana (McKiernan et ai., 2003; Singh ja Fawcett, 2008). Näissä tutkimuksissa tutkittiin deaktivoitumista tehtävän suorittamisen aikana. Tuloksemme viittaavat siihen, että tämä skaalautuminen voi tapahtua jo tehtävän ennakointivaiheen aikana. Weissman et al. (2006) tutkivat yksilöllisten reaktioaikojen vaihtelua huomiokatkosten tunnistamiseksi. Tarkkaavaisuuden lupsahdukset voisivat liittyä vähentyneeseen DMN-deaktivaatioon. Vaikka (Weissman et al., 2006) tarkasteli tarkkaavaisuuden tason luonnollista vaihtelua, meidän tuloksemme osoittavat, että useiden DMN-alueiden deaktivaatiota voidaan myös virittää ylhäältä alaspäin ennen tehtävän ärsykkeen esittämistä.
Syvällisimmät viritysvaikutukset havaittiin vasemmanpuoleisilla mediaalisilla prefrontaalisilla, temporaalisilla ja visuaalisilla alueilla. Seuraavassa keskustelemme näiden vaikutusten mahdollisista vaikutuksista näille alueille suhteessa niiden ehdotettuun tehtävään.
Mediaalisen prefrontaalisen aivokuoren aktiivisuus on aiemmin liitetty mielen vaelteluun (Bertossi et ai., 2017), muistin hakemiseen (Euston et ai., 2012) ja sydän- ja verenkiertoelimistön hallintaan (Wong et ai., 2007). Tämä mediaalinen prefrontaalinen deaktivaatio viittaa siihen, että jokin näistä toiminnoista tukahdutetaan jo kognitiivisen haasteen ennakoinnissa.
Deaktivaation virittäminen odotettavissa olevaan vaikeuteen visuaalisilla ja temporaalisilla alueilla voi liittyä epäolennaisen visuaalisen ja auditiivisen aistitulon tukahduttamiseen. Virittyminen kahdenvälisillä temporaalialueilla voi erityisesti liittyä magneettikuvauslaitteen tuottaman melun tukahduttamiseen. On kohtuullista olettaa, että äänen taso on samanlainen eri olosuhteissa. Se, että deaktivoitumisen taso on voimakkaampi vaikeiden vihjeiden kohdalla, viittaa siihen, että ympäristön äänen käsittely tukahdutetaan voimakkaammin, jos odotetaan vaikeaa tehtävää. Deaktivoitumista visuaalisissa aivokuorialueissa on raportoitu harvemmin kuin deaktivoitumista temporaalisilla alueilla. Smith ym. (2000) ovat kuitenkin aiemmin raportoineet yhteyden laajalle levinneen deaktivaation välillä näköaivokuorella ja huomion kiinnittämisen tiettyyn kuvan osaan. Giesbrecht et al. (2006) raportoivat myös visuaalisen aktivaation modulaatiota, joka liittyy peiteltyyn huomion kiinnittämiseen näkökentän keskipisteen lähellä oleviin kohteisiin verrattuna perifeerisen näkökentän kohteisiin. Nämä havainnot osoittavat, että visuaalisen aivokuoren aktivaatiota voidaan moduloida joko foveaalisiin tai perifeerisiin alueisiin kohdistuvalla salatulla huomiolla. Tutkimuksemme lisää uuden ulottuvuuden näihin havaintoihin, sillä se viittaa siihen, että aktivoitumista visuaalisissa aivokuoreissa ei moduloi ainoastaan peitellyn visuaalisen huomion sijainti vaan myös visuaalisen haasteen odotettu vaikeus; ja koska vihjeet olivat sijainniltaan ja kooltaan identtisiä, tätä prosessia näyttäisi säädeltävän ylhäältä alaspäin suuntautuvalla tavalla.
Tunnistimme myös joukon aivoalueita, jotka osoittivat lisääntynyttä aktivoitumista kognitiivisen haasteen ennakoinnissa. Tämä verkko osoitti samankaltaisuutta keskeisen toimeenpanoverkon kanssa, joka on aiemmin liitetty työmuistitehtävien suorittamiseen (Lawrence et ai., 2003). Tämän verkon aktivoituminen ei kuitenkaan ollut viritetty odotetun tehtävän vaikeuden mukaan. Tämä viittaa siihen, että nämä alueet edustavat prosesseja, jotka ovat samanlaisia vaikeassa ja helpossa tilassa. Mahdollisesti tämän verkon aktiivisuus liittyy vihjeen arviointiin tai tehtäväkontekstin ja ohjeiden hakemiseen ja ylläpitämiseen.
On otettava huomioon joitakin rajoituksia, jotka voivat vaikuttaa tulostemme tulkintaan. Ensinnäkin emme sisällyttäneet tutkimuksia, joissa oli ärsykkeitä, joita ei edeltänyt vihje, joten ei ole mahdollista tutkia vihjeen vaikutusta kognitiiviseen suorituskykyyn. Aikarajoituksen vuoksi ei ollut mahdollista toistaa tehtäväämme käyttämällä identtisiä ärsykkeitä, mutta ilman vihjeitä.
Toiseksi käytimme huutomerkkiä osana vaikeaa kognitiivista haastetta osoittavaa vihjettä, koska tätä symbolia käytetään tyypillisesti, jos on tarve lisätä tarkkaavaisuutta. On mahdollista, että huutomerkillä itsellään on ollut merkitystä erilaisten aktivoitumismallien syntymiseen helpossa ja vaikeassa tilanteessa sen luontaisen merkityksen vuoksi. Tämä ei kuitenkaan vaikuta tulostemme päätulkintaan, paitsi että päätuloksemme voidaan luokitella alhaalta ylöspäin suuntautuvaksi reaktioksi ylhäältä alaspäin suuntautuvan reaktion sijasta. Lopuksi, koesuunnitelmamme ei anna meille mahdollisuutta tutkia, voidaanko faasiseen valppauteen liittyvä aktiivisuus yleistää muille alueille vai onko se aluekohtaista, koska se sisälsi vain visuaalis-havainnollisen kognitiivisen haasteen.
Johtopäätöksenä voimme todeta, että tässä tutkimuksessa olemme osoittaneet, että faasisen valppauteen virittyminen edustaa deaktivoitumisen tasoa useilla alueilla. Tämä vaikutus on voimakkain mediaalisessa prefrontaalisessa, visuaalisessa ja temporaalisessa aivokuoressa, mikä voi heijastaa kardiovaginaalisen kontrollin tukahduttamista, visuaalista prosessointia perifeerisessä näkökentässä ja magneettikuvausskannerin kohinan tukahduttamista. Nämä tulokset viittaavat siihen, että kognitiivista suorituskykyä helpottaa aivojen tila, joka on tiukasti sidoksissa odotuksiin tulevan kognitiivisen haasteen vaikeudesta.
Tietojen saatavuus
Raaka-aineisto tuotettiin Elisabeth-Tweestedenin sairaalassa. Johdetut tiedot, jotka tukevat tämän tutkimuksen tuloksia, ovat saatavissa vastaavalta kirjoittajalta perustellusta pyynnöstä.
Eettinen lausunto
Tämä tutkimus toteutettiin Alankomaiden sosiaalitukilain suositusten mukaisesti, lääketieteellinen eettinen komitea Brabant. Kaikki koehenkilöt antoivat kirjallisen tietoon perustuvan suostumuksen Helsingin julistuksen mukaisesti. Lääketieteellinen eettinen komitea Brabant hyväksyi protokollan.
Author Contributions
MD ja JJ kirjoittivat pääkäsikirjoituksen ja valmistelivat luvut. MD ja IS suorittivat kokeet. G-JR, NR ja JJ valvoivat työtä yhdessä. Kaikki kirjoittajat tarkistivat käsikirjoituksen.
Rahoitus
Tämän tutkimuksen on rahoittanut Alankomaiden kansallinen organisaatio ZonMW (Netherlands Organization for Health Research and Development, ZONMW; projektinumero: 842003004), projektin nimi: Clinical implementation of advanced MRI techniques for localization and monitoring of sensorimotor and cognitive functions in patients with brain tumors.
Conflict of Interest Statement
Tekijät ilmoittavat, että tutkimus suoritettiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Acknowledgments
Haluaisimme kiittää röntgenhoitajia William Pigmansia ja Maikel Brandsia heidän myötävaikutuksestaan tähän tutkimukseen.
Lyhenteet
DMN, oletustilaverkko; M, mies; F, nainen; GLM, yleinen lineaarinen malli; SPM, tilastollinen parametrinen kartoitus; CueE, helppoa ärsykettä osoittava vihje, jota ei seuraa ärsyke; CueD, vaikeaa ärsykettä osoittava vihje, jota ei seuraa ärsyke; StimE, helppoon ärsykkeeseen viittaava vihje, jota seuraa helppo ärsyke; StimD, vaikeaan ärsykkeeseen viittaava vihje, jota seuraa vaikea ärsyke; MRI, magneettikuvaus; ROI, kiinnostava alue; GDL, GNU datakieli; TIA, tehtävästä johtuva aktivaatioverkko; TID, tehtävästä johtuva deaktivointiverkko; SEM, keskivirhe.
Footnotes
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M., ja Smith, A. T. (2011). Huomion modulaatio visuaalisessa aivokuoressa muuttuu havainto-oppimisen aikana. Neuropsychologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi, E., Peccenini, L., Solmi, A., Avenanti, A. ja Ciaramelli, E. (2017). Mediaalisen prefrontaalisen aivokuoren transkraniaalinen tasavirtastimulaatio vaimentaa mielen vaeltelua miehillä. Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., ja Schacter, D. L. (2008). Aivojen oletusverkko: anatomia, toiminta ja merkitys sairauksien kannalta. Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Buracas, G. T., ja Boynton, G. M. (2002). Tapahtumiin liittyvien fMRI-kokeiden tehokas suunnittelu M-sekvenssien avulla. Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | CrossRef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P. ja Bushnell, M. C. (2015). Tarvitaanko reagoivaa oletustilaverkkoa onnistuneeseen työmuistitehtävien suorittamiseen? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., ja Paz-Alonso, P. M. (2016). Vaiheisen hälytyksen ja tietoisuuden väliset vuorovaikutukset fronto-striatal-verkossa. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | CrossRef Full Text | Google Scholar
Desmond, J. E., and Glover, G. H. (2002). Otoskoon arviointi toiminnallisen magneettikuvauksen (fMRI) neurokuvantamistutkimuksissa: tilastolliset tehoanalyysit. J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., ja McNaughton, B. L. (2012). Mediaalisen prefrontaalisen aivokuoren rooli muistissa ja päätöksenteossa. Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., ja Posner, M. I. (2005). Huomioverkkojen aktivoituminen. Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T., Raz, A. ja Posner, M. I. (2002). Huomioverkkojen tehokkuuden ja riippumattomuuden testaaminen. J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/089892902317361886
PubMed Abstract | CrossRef Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G. ja Mangun, G. R. (2006). Pre-kohteen aktiivisuus visuaalisessa aivokuoressa ennustaa käyttäytymissuoritusta spatiaalisissa ja ominaisuuksien huomiotehtävissä. Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009). Vapaaehtoisen reaktion nopeuttaminen varoitussignaalilla. Psychophysiology 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R., Sorsa, M., and Siivonen, P. (2008). 6/6- ja 4/8-vuorojärjestelmien vaikutukset komentosiltaupseerien uneliaisuuteen. Chronobiol. Int. 25, 413-423. doi: 10.1080/07420520802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A. ja Buckner, R. L. (2012). Epäonnistuminen huomiokontrollin moduloinnissa pitkälle edenneessä ikääntymisessä liittyy valkoisen aineen patologiaan. Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., and Duyn, J. H. (2007). fMRI-tutkimus ponnisteluista ja tiedonkäsittelystä työmuistitehtävässä. Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., and Rutten, G. J. (2017). ”Ehdotus järjestelmäksi, joka helpottaa fMRI-tutkimusten kvantitatiivista toistamista”, in Proceedings of the Human Brain Mapping Conference 2017 June 25-29, Vancouver, Canada. Saatavilla verkossa osoitteessa: https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., and Baker, C. I. (2009). Ympyräanalyysi systeemien neurotieteessä: kaksoissukelluksen vaarat. Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A., and Bartolomeo, P. (2011). Faasinen auditiivinen hälytys parantaa visuaalista tietoista havaitsemista. Conscious. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G. ja Stein, E. S. (2003). Useat hermoverkot välittävät jatkuvaa huomiota. J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/089892903770007416
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, G., Sobering, G., Duyn, J. ja Moonen, C. T. (1993). Toiminnallinen magneettikuvaustekniikka, jossa yhdistyvät kaikusiirtymisen periaatteet ja havaintojakso (PRESTO). Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | CrossRef Full Text | Google Scholar
Macleod, J. W., Lawrence, M. A., McConnell, M. M., Eskes, G. A., Klein, R. M. ja Shore, D. I. (2010). ANT:n arviointi: psykometrisiä ja teoreettisia näkökohtia huomioverkkotestistä. Neuropsychology 24, 637-651. doi: 10.1037/a0019803
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., D’Angelo, B. R. D., Kaufman, J. N. ja Binder, J. R. (2006). ”Tajunnanvirran” keskeyttäminen: fMRI-tutkimus. Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J. K. ja Binder, J. R. (2003). Tehtävän aiheuttamaan deaktivoitumiseen vaikuttavien tekijöiden parametrinen manipulointi toiminnallisessa neurokuvantamisessa. J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/089892903321593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R. D., ja Brady, C. B. (1993). Faasinen ja tooninen vireystila Alzheimerin taudissa. Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., and Ramsey, N. F. (2008). Parannettu herkkyys nopealla kolmiulotteisella veren happipitoisuudesta riippuvaisella toiminnallisella magneettikuvauksella: SENSE-PRESTOn ja 2D-EPI:n vertailu 3 T:ssä. NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., and de Marco, G. (2010). Valppaus nuorilla terveillä koehenkilöillä: fMRI-tutkimus aivojen alueiden vuorovaikutteisuudesta, jota varoitussignaali lisää. Brain Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H., et al. (2009). Työmuistin kuormituksen vaikutus FMRI-lepotilan malliin myöhemmissä lepovaiheissa. PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E. (2015). Aivojen oletustilaverkko. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E., MacLeod, J. W., Snyder, A. Z., Powers, W. J., Gusnard, D. A. ja Shulman, G. L. (2001). Aivotoiminnan oletustila. Proc. Natl. Acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Petersen, S. E., et al. (1999). Alueet, jotka osallistuvat liikkuvien kohteiden suuntaodotusten koodaamiseen ja soveltamiseen. J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K. D., ja Fawcett, I. P. (2008). Ihmisen oletustilaverkon ohimenevä ja lineaarisesti porrastettu deaktivointi visuaalisen havaintotehtävän avulla. Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, A. T., Singh, K. D., and Greenlee, M. W. (2000). Ihmisen näköaivokuoren aktiivisuuden tarkkaavaisuuden tukahduttaminen (Attentional suppression of activity in the human visual cortex). Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., de Simone, A., Krause, B. J., Specht, K., Hessemann, V., Radermacher, I., et al. (1999). Sisäisen vireystilan toiminnallinen anatomia: todisteet oikean aivopuoliskon froto-parietal-talamus-aivorunkoverkosta. Neuropsychologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., ja Willmes, K. (2001). Intrinsisen ja faasisen vireystilan toiminnallisesta neuroanatomiasta. Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | CrossRef Full Text | Google Scholar
Thiel, C. M., Zilles, K., ja Fink, G. R. (2004). Aivokorrelaatit visuospatiaalisen huomion hälyttämisestä, suuntaamisesta ja uudelleen suuntaamisesta: tapahtumiin liittyvä fMRI-tutkimus. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Aktivointien automatisoitu anatominen merkitseminen SPM:ssä käyttäen MNI MRI yhden koehenkilön aivojen makroskooppista anatomista palstoitusta. Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | CrossRef Full Text | Google Scholar
van Gelderen, P., Ramsey, N. F., Liu, G., Duyn, J. H., Frank, J. A., Weinberger, D. R., et al. (1995). Ihmisen aivojen kolmiulotteinen toiminnallinen magneettikuvaus kliinisellä 1,5-T-skannerilla. Proc. Natl. Acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N., and Henik, A. (2011). Faasinen valppaus voi moduloida toimeenpanovalvontaa tehostamalla visuaalisten ärsykkeiden globaalia käsittelyä. Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., Visscher, K. M., and Woldorff, M. G. (2006). Hetkellisten huomiokatkosten hermostolliset perusteet. Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | CrossRef Full Text | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S. ja Shoemaker, J. K. (2007). Ventraalinen mediaalinen prefrontaalinen aivokuori ja kardiovaginaalinen kontrolli tajuissaan olevilla ihmisillä. Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka, H. T., Saito, D. N., Uchiyama, Y. ja Sadato, N. (2010). Faasisen vireystilan hermosubstraatit: toiminnallinen magneettikuvaustutkimus. Neurosci. Res. 68, 51-58. doi: 10.1016/j.neures.2010.05.005
PubMed Abstract | CrossRef Full Text | Google Scholar