Introduction
Pesquisas anteriores mostram que as pessoas podem usar uma deixa para se prepararem mentalmente para um desafio cognitivo. Se os participantes são instruídos a prepararem-se mentalmente no momento em que uma taco é apresentada, isto foi definido como alerta de fase. A capacidade de influenciar o nosso nível de alerta pode ser importante, especificamente se o alerta também tiver um custo. Se a demanda de alerta for muito alta, isso pode potencialmente levar à fadiga (Härmä et al., 2008) ou problemas relacionados ao estresse. Neste estudo, o objetivo é examinar se e como adultos saudáveis sintonizam seu nível de alerta com a dificuldade esperada de uma tarefa cognitiva futura.
Se uma sugestão for apresentada imediatamente antes de um estímulo de tarefa, as pessoas geralmente são capazes de responder mais rapidamente em tarefas de resposta motora simples (Fan et al., 2002; Macleod et al., 2010; Weinbach e Henik, 2011) e perceber estímulos visuais degradados com mais precisão (Kusnir et al., 2011). Estudos de imagem têm mostrado que o alerta de fase está associado ao aumento da ativação nas regiões frontal, parietal, talâmica (Shulman et al., 1999; Fan et al., 2005; Yanaka et al., 2010), temporo-occipital (Thiel et al., 2004) e cerebral visual (Bartolucci e Smith, 2011). Estes estudos de imagem têm fornecido informações valiosas sobre a representação de alerta de fase no cérebro. No entanto, a atenção de fase tem sido geralmente estudada como um fenômeno on/off ou a análise tem sido restrita ao córtex visual. Portanto, não está claro se a vigilância também reflete a dificuldade esperada do desafio cognitivo em redes cerebrais inteiras.
Outros estudos anteriores indicaram que a desativação de regiões cerebrais que fazem parte da rede de modo padrão (DMN) pode facilitar a execução de tarefas cognitivas. Primeiro, estudos de execução de tarefa mostram que um nível crescente de dificuldade de tarefa está associado a uma desativação crescente (McKiernan et al., 2003, 2006; Jansma et al., 2007; Singh e Fawcett, 2008; Pyka et al., 2009; Hedden et al., 2012; Čeko et al., 2015). Em segundo lugar, Jansma et al. (2007) mostraram que a parte pré-frontal medial do DMN só sintoniza a dificuldade da tarefa, que o participante poderia prever, mas não foi afetado pela dificuldade real de cada estímulo, que o participante não poderia prever. Finalmente, Weissman et al. (2006) descobriram que a desativação na DMN pouco antes do estímulo era mais fraca se os sujeitos respondessem relativamente devagar. Talvez estes sujeitos tenham respondido mais lentamente porque estavam temporariamente menos alerta.
Para o nosso estudo, desenhamos uma tarefa de percepção visual com uma condição fácil e difícil. As sugestões forneceram informações sobre a dificuldade do próximo estímulo, mas não forneceram nenhuma informação que facilitasse a tarefa em si. Em metade das provas, a deixa não foi seguida por um estímulo. Somente estas tentativas foram analisadas para a atividade cerebral, de modo que pudemos isolar completamente a ativação associada à taco, da ativação associada à execução da tarefa. Hipótese é que os participantes sintonizem sua atividade com a dificuldade esperada da tarefa.
Materiais e Métodos
Participantes
Participantes foram recrutados através de anúncio online. Os participantes foram excluídos se relatassem um histórico de distúrbios neurológicos ou psiquiátricos significativos, ou contra-indicações para a ressonância magnética (ressonância magnética) (objetos metálicos dentro ou ao redor do corpo, claustrofobia ou gravidez). Este estudo foi realizado de acordo com as recomendações da Medical Research Involving Human Subjects Act (WMO), Medical Research Ethics Committee Brabant. O protocolo foi aprovado pelo Comitê de Ética em Pesquisa Médica (número do protocolo: NL51147.028.14). Todos os sujeitos deram consentimento informado por escrito de acordo com a Declaração de Helsinki.
Vinte e dois voluntários saudáveis destros participaram do estudo. Os dados de dois participantes foram excluídos devido a artefatos de scanner. Os resultados são relatados para os 20 participantes restantes .
Desenho da tarefa
Desenhamos uma tarefa (com desenho relacionado ao evento) que nos permitiu examinar os efeitos de antecipação relacionados à dificuldade esperada de uma tarefa, sem confundir os efeitos da execução da tarefa.
Estímulos da tarefa consistiram de nove setas em um layout de três por três em uma tela preta (Figura 1). Um “estímulo fácil” continha oito setas na direção correta e uma seta na direção oposta. Um “estímulo difícil” continha cinco setas na direção correta e quatro setas na direção oposta (Figura 1). Os participantes foram instruídos a pressionar um botão com a mão correspondente à direção da maioria das setas. Os estímulos fáceis e difíceis foram apresentados em uma ordem aleatória. Os estímulos de linha de base consistiram de uma tela preta estacionária com o texto: “você tem uma pausa de 30 s” em holandês e em letras brancas. Os estímulos foram apresentados pelo software “presentation”.
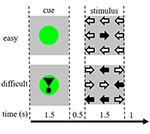
Figura 1. Representação dos estímulos visuais. Cada ensaio foi de 4,5 s. Em metade dos ensaios o taco não foi seguido de um estímulo. Estes ensaios foram utilizados para a análise da ressonância magnética funcional (fMRI).
Cada ensaio começou com um taco indicando a dificuldade do estímulo apresentado posteriormente. Um ponto verde indicou um “estímulo fácil”, um ponto verde com ponto de exclamação indicou um “estímulo difícil” (Figura 1).
Em metade das tentativas, a deixa não foi seguida por um estímulo, na outra metade o estímulo foi apresentado 500 ms após o desaparecimento da deixa. Isto resultou em quatro condições: (1) uma deixa para um estímulo fácil, não seguida por um estímulo (“CueE”); (2) uma deixa para um estímulo difícil, não seguida por um estímulo (“CueD”); (3) uma deixa para um estímulo fácil, seguida por um estímulo fácil (“StimE”); e (4) uma deixa para um estímulo difícil, seguida por um estímulo difícil (“StimD”). Todas as condições foram contrastadas com a linha de base. Os resultados da ressonância magnética funcional (fMRI) foram baseados em CueE e CueD. Os resultados de desempenho foram baseados no StimE e no StimD. CueE e CueD foram apresentados em ordem pseudo-aleatória usando sequências M para maximizar a independência do regressor do modelo linear geral (GLM) entre condições e otimizar a eficiência do projeto (Buracas e Boynton, 2002). Os participantes não puderam prever se um taco seria seguido ou não por um estímulo.
A duração de cada ensaio foi de 4.500 ms. As sugestões foram apresentadas em t = 0 para 1.500 ms e os estímulos foram apresentados em t = 2.000, também para 1.500 ms (Figura 1). A tarefa foi realizada em duas séries de 68 ensaios. A linha de base consistiu em três períodos de descanso de 30 s, antes, depois e entre as duas corridas. A duração total da tarefa foi de 11,7 minutos. Esta foi a segunda tarefa da sessão e começou após os participantes estarem deitados aproximadamente 10 min no scanner.
Os participantes foram instruídos a se prepararem para um estímulo fácil ou difícil com base nas informações da táctica (Figura 1) e a indicarem o mais precisa e rapidamente possível a direção da maioria das setas no estímulo. Os participantes praticaram a tarefa fora do scanner seguindo um protocolo de prática padrão de 3 minutos. Este protocolo foi repetido se o desempenho fosse insuficiente (abaixo de 70% de precisão para os estímulos difíceis).
Um espelho preso à bobina da cabeça permitiu aos participantes ver uma tela de projeção transparente posicionada atrás da cabeça. Um projetor de vídeo dentro da sala do scanner projetou os estímulos da tarefa na tela. Duas caixas de botões pneumáticos com pressão de ar foram usadas para gravar as respostas. Os participantes usaram o polegar esquerdo ou o indicador para responder “esquerda” e o direito para responder “direita”
Image Acquisition
Scans foram realizados em um scanner Philips Achieva 3T (Philips Medical Systems, Best, Holanda) usando uma bobina de cabeça SENSE de 32 canais. Uma imagem estrutural em T1 3D foi adquirida para fins de registro anatômico . imagens fMRI foram obtidas usando uma seqüência de pulsos PRESTO 3D . Seis digitalizações dummy foram adquiridas e depois descartadas pelo scanner.
Image Pre-processing
fMRI dados foram pré-processados usando mapeamento estatístico paramétrico (SPM12; Wellcome Trust Centre for Neuroimaging, University College London, London, UK2). As varreduras de uma sessão foram realinhadas para a primeira varredura para corrigir o movimento do sujeito usando uma abordagem de mínimos quadrados, uma transformação espacial de seis parâmetros (corpo rígido), e uma estimativa de 2° grau B-spline. As varreduras foram co-registradas para o T1 usando um modelo de corpo rígido. Os parâmetros foram estimados através da função de informação mútua normalizada. As imagens foram ressecadas por uma linha reta de 4° grau B. O T1 foi normalizado espacialmente em espaço MNI padrão usando uma regularização de polarização muito leve (0,0001) e uma linha B-Spline de 4º grau. Os parâmetros resultantes foram aplicados a todas as varreduras funcionais, a fim de minimizar as diferenças anatômicas e, portanto, permitir a análise em grupo. Finalmente, todas as varreduras foram suavizadas espacialmente com um filtro Gaussiano 3D (largura total na metade do máximo: 8 mm) para minimizar ainda mais o efeito das diferenças anatômicas funcionais.
Análise fMRI individual
Análise de regressão GLM relacionada a eventos foi realizada para cada voxel para gerar mapas de ativação individuais usando uma abordagem de massaunivariada com um modelo de autocorrelação aproximada global do AR(1) e um filtro de passagem alta (corte de 128 s). A linha de base não foi modelada explicitamente. A função básica era uma HRF canônica sem derivadas. Foi utilizado um limiar de mascaramento de 0,8. Foram utilizados regressores separados para cada condição (CueE, 17 timepoints; CueD, 16 timepoints; StimE, 17 timepoints; StimD, 18 timepoints; e um regressor de incómodo para períodos blanc na tarefa relacionada com a sequência m, 68 timepoints). Nenhum timepoints foi excluído da análise. Os mapas Beta foram transformados para refletir a mudança real de sinal percentual em cada voxel. Apresentamos apenas os resultados para os regressores CueE e CueD, reflectindo as alterações de sinal para tacos sem um estímulo. Beta e mapas t estatísticos foram verificados visualmente para artefatos principais.
Análise fMRI do grupo
Uma análise fMRI de segundo nível foi realizada com regressão linear em cada voxel para visualização e seleção da região de interesse (ROI). Uma análise de ROI foi realizada com a linguagem de dados GNU (GDL3), usando mapas individuais de mudança de sinal percentual do sujeito gerado pela análise GLM. Os voxels dentro de um ROI ou rede foram calculados como média para calcular a diferença de sinal entre CueE, CueD e baseline.
Seleção de ROI
Máximos e mínimos locais supra-tratoriais foram determinados para CueE e CueD por SPM com um limiar não corrigido (p < 0,001; Tabela 1). Os ROIs cúbicos de tamanho e forma predefinidos foram colocados sobre os máximos locais para o CueE-baseline e CueD-baseline contrastam separadamente dentro de um raster pré-definido (Jansma e Rutten, 2017; Tabela 2). Os ROIs foram incluídos na análise se continham um máximo local significativo para o CueE ou CueD. Ao definir os ROIs nos máximos locais para ambas as condições separadamente, evita-se um enviesamento para CueE ou CueD. Além disso, a forma e tamanho pré-definidos das ROIs dentro de um raster reduz o efeito da circularidade, porque as fronteiras destas ROIs não são afectadas pelo ruído (Kriegeskorte et al., 2009). Além disso, este método facilita a comparação quantitativa de resultados entre condições, e potencialmente entre diferentes estudos, facilitando assim a reprodutibilidade quantitativa dos resultados da fMRI. Colocação dos ROIs sobre os máximos locais de potência otimizada. No entanto, sabe-se que a localização do pico de atividade é afetada pelo ruído, portanto a localização exata das ROIs pode sempre se reproduzir para cada ROI. Isto não afecta os resultados da rede, uma vez que todos os ROIs são médios e a localização exacta, portanto, não é tão relevante.
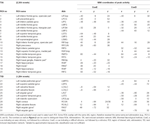
Table 1. Padrão de atividade.
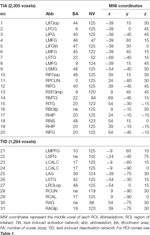
Tabela 2. Características da região de interesse (ROI).
ROIs sobre regiões com um aumento na mudança de sinal foram combinados em uma rede de ativação induzida por tarefa (“TIA”). Os ROIs sobre regiões com uma diminuição na mudança de sinal foram combinados em uma rede de desativação induzida por tarefa (“TID”; Tabelas 1, 2). Sempre que possível, os nomes das ROI foram determinados usando o atlas AAL (Tzourio-Mazoyer et al., 2002).
Tabela 2 e Figura 2 fornecem uma visão geral do tamanho e localização de todas as ROIs. Como o tamanho e os limites dos ROIs são predefinidos, é possível que a atividade média no ROI não seja significativamente diferente da linha de base, por exemplo, porque o número de voxels ativados dentro do ROI é relativamente pequeno. Embora a atividade média no nível da rede seja significativamente diferente da linha de base para todas as condições (Tabela 3), este não é o caso de oito dos 31 ROIs individuais para CueE e três dos 31 ROIs para CueD (Tabela 4). Apenas os voxels que continham sinal para cada participante foram incluídos na análise. O tamanho máximo de um ROI é 125 voxels, alguns ROIs incluem menos de 125 voxels porque estão posicionados perto do crânio.
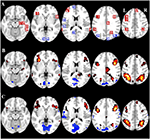
Figura 2. Visão geral da localização da região de interesse (ROI) e resultados de fMRI do cérebro inteiro. (A) ROIs para ativação induzida por tarefa (vermelho) e desativação induzida por tarefa (azul) nota: ROI 22 não é exibido porque a localização está fora das fatias escolhidas. As coordenadas MNI, número BA e tamanho das ROIs podem ser encontrados na Tabela 2. (B) Representação visual dos valores t para CueE vs. linha de base (≥3 = vermelho para amarelo, ≤ -3 = azul para azul claro), (C) Valores t para CueD vs. linha de base (≥3 = vermelho para amarelo, ≤ -3 = azul para azul claro). As imagens estão em orientação neurológica (L = esquerda, R = direita), nomes, abreviações, coordenadas MNI dos picos de ativação voxel podem ser encontrados na Tabela 1.

Tabela 3. Resultados da rede.
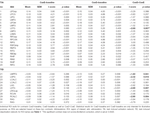
Tabela 4. Resultados do ROI.
Testes com Hipóteses
Testes com as seguintes hipóteses neste estudo:
Hipótese 1: A TIA mostrará aumento da mudança de sinal para CueD, em comparação com CueE.
Hipótese 2: A TID mostrará menor mudança de sinal para CueD, comparado ao CueE.
Análise estatística
Análise de potência não foi realizada, pois foi a primeira vez que este desenho de tarefa específica foi apresentado aos participantes. Portanto não há conhecimento da porcentagem esperada de mudança de sinal e variabilidade das condições (Desmond e Glover, 2002).
A precisão da tarefa foi calculada para StimE e StimD como a porcentagem de respostas corretas. Os tempos de reacção foram calculados para o StimE e StimD sobre todas as respostas correctas.
Uma amostra de testes t foi usada para comparar a variação percentual do sinal para TIA e TID em relação à linha de base para CueE e CueD. Testes t de amostra pareados de um lado foram usados para comparar a mudança de sinal, precisão e diferenças de tempo de reação entre CueD e CueE (Hipóteses 1 e 2).
Testes pós-hocais envolveram testes t separados para cada ROI dentro de cada rede. A análise do ROI foi corrigida em Bonferroni se o teste omnibus da rede não foi significativo. Apresentamos valores de p não corrigidos e um comentário declarando se este valor de p teria sobrevivido à correção de Bonferroni para todas as comparações múltiplas. SPSS 24 foi usado para todas as análises estatísticas.
Resultados
Performance
A precisão para os estímulos fáceis (“StimE”) foi de 98% ± 1,2% SEM e para os estímulos difíceis (“StimD”) 85% ± 2,6%. O tempo de reação para StimE foi de 682 ± 20 ms e para StimD 1.138 ± 27 ms. Ambas as condições mostram taxas de precisão suficientes para se ter a certeza de que a tarefa foi executada conforme instruído por todos os participantes. Como hipótese, os participantes responderam menos precisos (T = -4,3, p < 0,001) e mais lentos (T = 17,8, p < 0,001) para o StimD do que para o StimE (Figura 3), indicando que o contraste entre os estímulos de fácil e dificuldade foi bem sucedido.
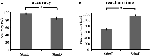
Figura 3. Resultados de desempenho. (A) Precisão para ambas as categorias de estímulos, (B) tempo de reação para ambas as categorias de estímulos. As barras de erro representam o erro padrão da média (SEM). *p ≤ 0.001. Abreviaturas: StimE, um taco indicando um estímulo fácil, seguido por um estímulo fácil; StimD, um taco indicando um estímulo difícil, seguido por um estímulo difícil.
Ressonância magnética funcional
Descritivos
A rede TIA consistiu de 20 ROIs, dos quais 10 foram baseados em máximos locais para CueE, cinco foram baseados em máximos locais para CueD, e cinco continham um CueE e um máximo CueD (Tabela 1). Destes 20 ROIs, 19 apresentaram uma actividade significativa em comparação com a linha de base para CueE, 18 para CueD (Tabela 1). A variação média de sinal na TIA em comparação com a linha de base para CueE foi 0,12 ± 0,02 (t = 6,11, p < 0,001) e para CueD 0,12 ± 0,02 (t = 7,61, p < 0,001).
A rede TID consistiu em 11 ROIs, dos quais três foram baseados em máximos locais para CueE, seis foram baseados em máximos locais para CueD, e dois continham um CueE e um CueD máximo (Tabela 1). Destas 11 ROIs, quatro apresentaram actividade significativa em relação à linha de base para CueE, 10 para CueD (Tabela 1). A variação média do sinal na TID em comparação com a linha de base para CueE foi -0,07 ± 0,02 (t = 2,68, p < 0,01), e para CueD -0,12 ± 0,02 (t = 5,37, p < 0,001; Tabela 3).
Ajustamento à Dificuldade Esperada
O aumento do sinal na TIA não foi significativamente mais forte para CueD comparado com CueE (t = -0,24, p = 0,41) indicando que o nível de ativação da TIA não está ajustado à dificuldade esperada (Figura 4, Tabela 3). A análise do ROI pós- hoc não mostrou nenhum dos 20 ROIs dentro do TIA com um aumento significativo do sinal para CueD comparado ao CueE após a aplicação da correção de Bonferroni (Tabela 4, Figura 5).
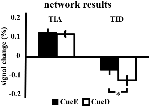
Figura 4. Visão geral dos resultados da rede. As barras de erro representam SEM. *p ≤ 0.05 para taco difícil vs. taco fácil. Abreviaturas: TIA, rede de ativação induzida por tarefa; TID, rede de desativação induzida por tarefa; CueE, um taco indicando um estímulo fácil, não seguido por um estímulo; CueD, um taco indicando um estímulo difícil, não seguido por um estímulo.
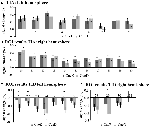
Figure 5. Visão geral das respostas individuais ao ROI dentro da rede de ativação induzida por tarefa, hemisfério esquerdo (A) e direito (B), e desativação induzida por tarefa, hemisfério esquerdo (C) e direito (D). As barras de erro representam SEM. *p ≤ 0,05 para CueD vs. CueE, ver Figura 2 para a localização das ROIs, ver Tabela 1 para os nomes, abreviaturas e coordenadas MNI das ROIs.
A diminuição do sinal na TID foi significativamente mais forte para CueD em comparação com CueE (T = -2,48, p = 0,01) indicando que o nível da TID está sintonizado com a dificuldade esperada (Tabela 3, Figura 4). A análise do ROI pós- hoc mostrou quatro das 11 ROIs da TID com uma diminuição significativa do sinal para o CueD em comparação com o CueE, nomeadamente LMPFG (T = -1,83, p = 0,04), LSPG (T = -2,32, p = 0,02), LSTG (T = -2,69, p < 0,01), e RCALC (T = -2,90, p < 0,01). Estes resultados indicam que o ajuste à dificuldade esperada é mais forte nas ROIs frontal, temporal e visual esquerda (Tabela 4, Figura 5).
Discussão
Neste estudo, examinamos se e como a atenção de fase está ajustada à dificuldade esperada de um desafio cognitivo que se aproxima. A atenção fásica foi refletida tanto nas regiões do cérebro ativadas quanto nas desativadas. A rede ativada incluiu regiões da rede executiva central. A rede desativada incluiu regiões da DMN, bem como cortices visuais. O nível de desativação escalou com a dificuldade esperada do próximo estímulo, enquanto o nível de ativação não o fez. Estes resultados sugerem que a modulação de alerta de fase é predominantemente refletida em processos de supressão gradual que são irrelevantes para a tarefa atual (por exemplo, processos cardio-vagal, auditivos e visuais). Possivelmente, isso ocorreu porque esses processos poderiam interferir nos próximos desafios cognitivos.
Estudos de imagem anteriores de alerta de fase utilizaram um nível de dificuldade, apenas apresentaram regiões cerebrais ativadas (Shulman et al., 1999; Sturm e Willmes, 2001; Thiel et al., 2004; Fan et al., 2005; Périn et al., 2010; Yanaka et al., 2010), ou focaram apenas a atividade no córtex visual (Bartolucci e Smith, 2011). Estes estudos identificaram uma rede fronto-parietalâmica, na sua maioria, lateralizada à direita, por vezes combinada com regiões corticais motoras e visuais, que foi activada por um taco. Existem diferenças substanciais nas redes ativadas entre os estudos. Algumas das diferenças entre as redes em estudos anteriores podem ser explicadas pela tarefa que se seguiu à táctica. Por exemplo, Thiel et al. (2004) apenas identificaram a ativação em regiões corticais visuais usando uma tarefa de percepção visual, enquanto Fan et al. (2005) e Yanaka et al. (2010) identificaram a ativação talâmica e motora com uma tarefa de resposta motora. Embora Bartolucci e Smith (2011) apresentem um desenho elegante de tarefa com quatro níveis de dificuldade de uma tarefa de discriminação de orientação, eles apresentaram apenas a ativação no córtex visual. Portanto, não está claro como outras regiões cerebrais responderam a esta tarefa. A rede de ativação induzida por tarefa, identificada no estudo atual é mais semelhante à rede encontrada por Shulman et al. (1999), que usou um tipo similar de tarefa de processamento visual.
Não está claro se a ativação de fase é uma resposta de baixo para cima ou de cima para baixo a uma taco não-informativa (Thiel et al., 2004; Hackley, 2009; Périn et al., 2010; Bartolucci e Smith, 2011; Chica et al., 2016). Phasic alertness has been considered by some as bottom-up (Sturm et al., 1999; Hackley, 2009), by others as top-down (Bartolucci e Smith, 2011), and by others as a combination (Thiel et al., 2004; Périn et al., 2010) response. Argumentamos que é mais adequado rotular a resposta do cérebro a uma taco como uma combinação de processamento bottom-up e top-down porque os participantes respondem a uma taco externa e a uma instrução associada a esta taco. Contudo, nossos principais resultados dizem respeito à diferença na resposta a duas dicas similares indicando um tipo diferente de estímulo de tarefa. Portanto, argumentamos que a diferença resultante entre a deixa indicando uma tarefa difícil e a deixa indicando uma tarefa fácil pode representar um nível de controle top-down relacionado à interpretação da deixa e da instrução associada.
A rede de desativação induzida pela tarefa incluiu regiões cerebrais do DMN, a saber, o giro pré-frontal medial, gyri temporal superior e gyri angular (Shulman et al., 1999; Raichle et al., 2001; Raichle, 2015). É comumente levantada a hipótese de que a desativação da DMN auxilia no desempenho de tarefas, reduzindo processos internos que interferem com os desafios cognitivos (Raichle et al., 2001; Buckner et al., 2008; Raichle, 2015). Esta visão é apoiada por estudos que mostram que o nível de desativação da DMN é escalonado com a dificuldade de uma tarefa durante a execução da mesma (McKiernan et al., 2003; Singh e Fawcett, 2008). Estes estudos examinaram a desativação durante a execução da tarefa. Nossos resultados sugerem que essa escala já pode ocorrer durante a fase de antecipação da tarefa. Weissman et al. (2006) examinaram a variabilidade nos tempos de reação individual para identificar lapsos de atenção. Os lapsos de atenção podem estar associados com a redução da desativação da DMN. Enquanto (Weissman et al., 2006) examinaram a variação natural do nível de atenção, nossos resultados indicam que a desativação em várias regiões de DMN também pode ser sintonizada de cima para baixo antes que o estímulo da tarefa seja apresentado.
Os efeitos de sintonia mais profundos foram encontrados nas regiões pré-frontal, temporal e visual medial esquerda. Abaixo discutimos possíveis implicações desses efeitos para essas regiões em relação à sua função proposta.
Atividade do córtex pré-frontal medial tem sido previamente associada com o desvio da mente (Bertossi et al., 2017), recuperação de memória (Euston et al., 2012) e controle cardiovagal (Wong et al., 2007). Esta desativação pré-frontal medial sugere que qualquer uma destas funções já está suprimida em antecipação a um desafio cognitivo.
A sintonia da desativação com a dificuldade esperada nas regiões visuais e temporais pode estar relacionada à supressão da entrada sensorial visual e auditiva irrelevante. A sintonia nas regiões temporais bilaterais pode estar especificamente relacionada à supressão do ruído gerado pelo scanner de ressonância magnética. É razoável esperar que o nível de som seja semelhante entre as condições. O facto do nível de desactivação ser mais forte para as pistas difíceis sugere que o processamento do som ambiente é mais fortemente suprimido se for esperada uma tarefa difícil. A desativação nos cortices visuais é menos comumente relatada do que a desativação nas regiões temporais. Entretanto, Smith et al. (2000) relataram anteriormente uma associação entre a desativação generalizada no córtex visual e a atenção a uma parte específica de uma imagem. Giesbrecht et al. (2006) também relataram modulação da ativação visual associada à atenção oculta a objetos próximos ao centro de seu campo visual em relação a objetos no campo visual periférico. Esses achados indicam que a ativação nos cortices visuais pode ser modulada pela atenção encoberta a regiões foveais ou periféricas. Nosso estudo acrescenta uma nova dimensão a esses achados, pois sugere que a ativação nos cortices visuais não é modulada apenas pela localização da atenção visual oculta, mas também pela dificuldade esperada de um desafio visual; e como as pistas eram idênticas em localização e tamanho, esse processo parece regulado de cima para baixo.
Identificamos também um conjunto de regiões cerebrais que mostraram maior ativação em antecipação a um desafio cognitivo. Esta rede mostrou semelhança com a rede executiva central que foi anteriormente associada à execução de tarefas de memória de trabalho (Lawrence et al., 2003). No entanto, a ativação nesta rede não foi ajustada à dificuldade esperada da tarefa. Isto sugere que estas regiões representam processos que são similares para a condição difícil e fácil. Possivelmente, a atividade nesta rede está associada à avaliação do taco, ou recuperação e manutenção do contexto da tarefa e das instruções.
Somas limitações precisam ser levadas em consideração que podem afetar a interpretação dos nossos resultados. Em primeiro lugar, não incluímos ensaios com estímulos que não foram precedidos por uma taco, portanto não é possível examinar o efeito da taco no desempenho cognitivo. Devido à restrição de tempo, não foi possível repetir a nossa tarefa usando estímulos idênticos, mas sem taco.
Segundo, usamos um ponto de exclamação como parte do taco indicando um desafio cognitivo difícil, porque este símbolo é tipicamente usado se houver necessidade de aumentar a atenção. É possível que o ponto de exclamação em si tenha desempenhado um papel nos diferentes padrões de ativação para a condição fácil e difícil, devido ao seu significado inerente. Isto, no entanto, não afecta a interpretação principal dos nossos resultados, excepto que os nossos principais resultados podem ser classificados como uma resposta de baixo para cima em vez de uma resposta de cima para baixo. Finalmente, o nosso desenho experimental não nos permite examinar se a actividade relacionada com a alerteza de fase pode ser generalizada a outros domínios, ou se é específica a um domínio, uma vez que apenas inclui um desafio cognitivo visuo-perceptual.
Para concluir, neste estudo mostramos que a sintonia da alerteza de fase é representada pelo nível de desactivação em várias regiões. Este efeito é mais forte no córtex pré-frontal medial, visual e temporal que pode refletir uma supressão do controle cardiovagal, processamento visual no campo visual periférico e supressão do ruído do scanner de ressonância magnética. Estes resultados sugerem que o desempenho cognitivo é facilitado por um estado do cérebro que está firmemente acoplado às expectativas sobre a dificuldade de um próximo desafio cognitivo.
Dados Disponibilidade
Dados em bruto foram gerados no hospital Elisabeth-Tweesteden. Dados derivados que suportam os resultados deste estudo estão disponíveis do autor correspondente mediante solicitação razoável.
Declaração Ethics
Este estudo foi realizado de acordo com as recomendações da lei holandesa de apoio social, comitê de ética médica Brabant. Todos os sujeitos deram consentimento informado por escrito, de acordo com a Declaração de Helsinque. O protocolo foi aprovado pelo comitê de ética médica Brabant.
Contribuições dos autores
MD e JJ escreveram o manuscrito principal e prepararam as figuras. MD e IS realizaram os experimentos. G-JR, NR, e JJ supervisionaram conjuntamente o trabalho. Todos os autores revisaram o manuscrito.
Funding
Este estudo foi financiado pela ZonMW, uma organização nacional holandesa (Organização Holandesa de Pesquisa e Desenvolvimento em Saúde, ZONMW; Número do projeto: 842003004), título do projeto: Implementação clínica de técnicas avançadas de RM para localização e monitoramento das funções sensorimotoras e cognitivas em pacientes com tumores cerebrais.
Conflito de interesses
Os autores declaram que a pesquisa foi realizada na ausência de qualquer relação comercial ou financeira que pudesse ser interpretada como um potencial conflito de interesses.
Conhecimento
Gostaríamos de agradecer aos radiologistas William Pigmans e Maikel Brands pela sua contribuição para este estudo.
A abreviaturas
DMN, modo de rede padrão; M, masculino; F, feminino; GLM, modelo linear geral; SPM, mapeamento estatístico paramétrico; CueE, um taco indicando um estímulo fácil, não seguido por um estímulo; CueD, um taco indicando um estímulo difícil, não seguido por um estímulo; StimE, taco indicador de estímulo fácil, seguido de estímulo fácil; StimD, taco indicador de estímulo difícil, seguido de estímulo difícil; MRI, ressonância magnética; ROI, região de interesse; GDL, linguagem de dados GNU; TIA, rede de ativação induzida por tarefa; TID, rede de desativação induzida por tarefa; SEM, erro padrão da média.
Footnotes
- ^ https://www.zonmw.nl/nl/onderzoek-resultaten/kwaliteit-van-zorg/programmas/project-detail/topzorg/clinical-implementation-of-advanced-mri-techniques-for-localization-and-monitoring-of-sensorimotor-a/verslagen/
- ^ http://www.fil.ion.ucl.ac.uk/spm/software/spm12/
- ^ https://github.com/gnudatalanguage/gdl
Bartolucci, M., e Smith, A. T. (2011). A modulação atencional no córtex visual é modificada durante a aprendizagem perceptual. Neuropsicologia 49, 3898-3907. doi: 10.1016/j.neuropsychologia.2011.10.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Bertossi, E., Peccenini, L., Solmi, A., Avenanti, A., e Ciaramelli, E. (2017). A estimulação transcraniana por corrente contínua do córtex pré-frontal medial amortece o vaguear da mente nos homens. Sci. Rep. 7:16962. doi: 10.1038/s41598-017-17267-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Buckner, R. L., Andrews-Hanna, J. R., e Schacter, D. L. (2008). A rede padrão do cérebro: anatomia, função e relevância para a doença. Ann. N Y Acad. Sci. 1124, 1-38. doi: 10.1196/annals.1440.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Buracas, G. T., e Boynton, G. M. (2002). Projeto eficiente de experimentos de fMRI relacionados a eventos usando seqüências M. Neuroimage 16, 801-813. doi: 10.1006/nimg.2002.1116
PubMed Abstract | CrossRef Full Text | Google Scholar
Čeko, M., Gracely, J. L., Fitzcharles, M. A., Seminowicz, D. A., Schweinhardt, P., e Bushnell, M. C. (2015). É necessária uma rede de modo padrão de resposta para o desempenho bem sucedido de tarefas de memória de trabalho? J. Neurosci. 35, 11595-11605. doi: 10.1523/JNEUROSCI.0264-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Chica, A. B., Bayle, D. J., Botta, F., Bartolomeo, P., e Paz-Alonso, P. M. (2016). Interações entre o alerta de fase e a consciência na rede fronto-estriatal. Sci. Rep. 6:13. doi: 10.1038/srep31868
PubMed Abstract | CrossRef Full Text | Google Scholar
Desmond, J. E., e Glover, G. H. (2002). Estimando o tamanho da amostra em estudos de neuroimagem por ressonância magnética funcional (fMRI): análises de poder estatístico. J. Neurosci. Methods 118, 115-128. doi: 10.1016/s0165-0270(02)00121-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Euston, D. R., Gruber, A. J., and McNaughton, B. L. (2012). O papel do córtex pré-frontal medial na memória e na tomada de decisões. Neuron 76, 1057-1070. doi: 10.1016/j.neuron.2012.12.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., e Posner, M. I. (2005). A ativação de redes atencionais. Neuroimage 26, 471-479. doi: 10.1016/j.neuroimage.2005.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., e Posner, M. I. (2002). Testando a eficiência e independência das redes atencionais. J. Cogn. Neurosci. 14, 340-347. doi: 10.1162/089892902317361886
PubMed Abstract | CrossRef Full Text | Google Scholar
Giesbrecht, B., Weissman, D. H., Woldorff, M. G., e Mangun, G. R. (2006). Atividade pré-alvo em córtex visual prevê desempenho comportamental em tarefas espaciais e de atenção a características. Brain Res. 1080, 63-72. doi: 10.1016/j.brainres.2005.09.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Hackley, S. A. (2009). A aceleração da reacção voluntária através de um sinal de aviso. Psicofisiologia 46, 225-233. doi: 10.1111/j.1469-8986.2008.00716.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Härmä, M., Partinen, M., Repo, R., Sorsa, M., e Siivonen, P. (2008). Efeitos dos sistemas de relógio 6/6 e 4/8 na sonolência dos oficiais de ponte. Chronobiol. Int. 25, 413-423. doi: 10.1080/07420520802106769
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedden, T., Van Dijk, K. R., Shire, E. H., Sperling, R. A., Johnson, K. A., e Buckner, R. L. (2012). Falha em modular o controle atencional no envelhecimento avançado ligado à patologia da matéria branca. Cereb. Cortex 22, 1038-1051. doi: 10.1093/cercor/bhr172
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., and Duyn, J. H. (2007). fMRI estudo do esforço e processamento de informação numa tarefa de memória de trabalho. Hum. Brain Mapp. 27, 431-440. doi: 10.1002/hbm.20297
PubMed Abstract | CrossRef Full Text | Google Scholar
Jansma, J. M., e Rutten, G. J. (2017). “A proposal for a system that facilitates quantitative replication of fMRI studies”, em Proceedings of the Human Brain Mapping Conference 2017 25-29 de junho, Vancouver, Canadá. Disponível online em: https://archive.aievolution.com/2017/hbm1701/index.cfm?do=abs.viewAbs&abs=4047.
Google Scholar
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., e Baker, C. I. (2009). Análise circular em neurociência de sistemas: os perigos da dupla imersão. Nat. Neurosci. 12, 535-540. doi: 10.1038/nn.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Kusnir, F., Chica, A. B., Mitsumasu, M. A., e Bartolomeo, P. (2011). Alerta auditivo faseado melhora a percepção consciente visual. Consciente. Cogn. 20, 1201-1210. doi: 10.1016/j.concog.2011.01.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Lawrence, M. A., Ross, T. J., Hoffmann, H. G., e Stein, E. S. (2003). Múltiplas redes neurais intermediam a atenção sustentada. J. Cogn. Neurosci. 15, 1028-1038. doi: 10.1162/089892903770007416
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, G., Sobering, G., Duyn, J., and Moonen, C. T. (1993). Uma técnica de RM funcional que combina princípios de mudança de eco com um comboio de observações (PRESTO). Magn. Reson. Med. 30, 764-768. doi: 10.1002/mrm.1910300617
PubMed Abstract | CrossRef Full Text | Google Scholar
Macleod, J. W., Lawrence, M. A., McConnell, M. M., Eskes, G. A., Klein, R. M., and Shore, D. I. (2010). Appraising the ANT: psychometric and theoretical considerations of the attention network test. Neuropsicologia 24, 637-651. doi: 10.1037/a0019803
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., D’Angelo, B. R. D., Kaufman, J. N., e Binder, J. R. (2006). Interrompendo o “fluxo de consciência”: uma investigação fMRI. Neuroimage 29, 1185-1191. doi: 10.1016/j.neuroimage.2005.09.030
PubMed Abstract | CrossRef Full Text | Google Scholar
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J. K., and Binder, J. R. (2003). Uma manipulação paramétrica de fatores que afetam a desativação induzida por tarefas na neuroimagem funcional. J. Cogn. Neurosci. 15, 394-408. doi: 10.1162/089892903321593117
PubMed Abstract | CrossRef Full Text | Google Scholar
Nebes, R. D., e Brady, C. B. (1993). Phasic and tonic alertness in Alzheimer’s disease (Alzheimer). Cortex 29, 77-90. doi: 10.1016/s0010-9452(13)80213-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Neggers, S. F., Hermans, E. J., e Ramsey, N. F. (2008). Sensibilidade melhorada com ressonância magnética funcional rápida tridimensional dependente de oxigênio no sangue: comparação de SENSE-PRESTO e 2D-EPI a 3 T. NMR Biomed. 21, 663-676. doi: 10.1002/nbm.1235
PubMed Abstract | CrossRef Full Text | Google Scholar
Périn, B., Godefroy, O., Fall, S., and de Marco, G. (2010). Alertness in young healthy subjects: an fMRI study of brain region interactivity enhanced by a warning signal. Cérebro Cogn. 72, 271-281. doi: 10.1016/j.bandc.2009.09.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Pyka, M., Beckmann, C. F., Schöning, S., Hauke, S., Heider, D., Kugel, H., et al. (2009). Impacto da carga de memória de trabalho no padrão de estado de repouso FMRI em fases subsequentes de repouso. PLoS One 4:e7198. doi: 10.1371/journal.pone.0007198
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E. (2015). A rede de modo padrão do cérebro. Annu. Rev. Neurosci. 38, 433-447. doi: 10.1146/annurev-neuro-071013-014030
PubMed Abstract | CrossRef Full Text | Google Scholar
Raichle, M. E., MacLeod, J. W., Snyder, A. Z., Powers, W. J., Gusnard, D. A., e Shulman, G. L. (2001). Um modo padrão de funcionamento do cérebro. Proc. Natl. Acad. Sci. U S A 98, 676-682. doi: 10.1073/pnas.98.2.676
PubMed Abstract | CrossRef Full Text | Google Scholar
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E., Snyder, A. Z., Petersen, S. E., et al. (1999). Áreas envolvidas na codificação e aplicação de expectativas direcionais a objetos em movimento. J. Neurosci. 19, 9480-9496. doi: 10.1523/jneurosci.19-21-09480.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Singh, K. D., e Fawcett, I. P. (2008). Desativação transitória e linearmente graduada da rede de modo padrão humano através de uma tarefa de detecção visual. Neuroimage 41, 100-112. doi: 10.1016/j.neuroimage.2008.01.051
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, A. T., Singh, K. D., e Greenlee, M. W. (2000). Supressão atencional da atividade no córtex visual humano. Neuroreport 11, 271-277. doi: 10.1097/00001756-200002070-00010
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., de Simone, A., Krause, B. J., Specht, K., Hessemann, V., Radermacher, I., et al. (1999). Anatomia funcional do estado de alerta intrínseco: evidência de uma rede de tronco cerebral-froto-parietalâmica no hemisfério direito. Neuropsicologia 37, 797-805. doi: 10.1016/s0028-3932(98)00141-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Sturm, W., and Willmes, K. (2001). Sobre a neuroanatomia funcional de alerta intrínseco e de fase. Neuroimage 14, S76-S84. doi: 10.1006/nimg.2001.0839
PubMed Abstract | CrossRef Full Text | Google Scholar
Thiel, C. M., Zilles, K., and Fink, G. R. (2004). Correlatos cerebrais de alerta, orientação e reorientação da atenção visuoespacial: um estudo fMRI relacionado a eventos. Neuroimage 21, 318-328. doi: 10.1016/j.neuroimage.2003.08.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Rotulagem anatômica automatizada das ativações em SPM usando uma parcela anatômica macroscópica do cérebro mono-sujeito da ressonância magnética MNI. Neuroimage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | CrossRef Full Text | Google Scholar
van Gelderen, P., Ramsey, N. F., Liu, G., Duyn, J. H., Frank, J. A., Weinberger, D. R., et al. (1995). Ressonância magnética tridimensional funcional do cérebro humano em um scanner clínico 1.5-T. Proc. Natl. Acad. Sci. U S A 92, 6906-6910. doi: 10.1073/pnas.92.15.6906
PubMed Abstract | CrossRef Full Text | Google Scholar
Weinbach, N., e Henik, A. (2011). O alerta fasico pode modular o controle executivo, melhorando o processamento global dos estímulos visuais. Cognition 121, 454-458. doi: 10.1016/j.cognition.2011.08.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Weissman, D. H., Roberts, K. C., Visscher, K. M., e Woldorff, M. G. (2006). As bases neurais dos lapsos momentâneos em atenção. Nat. Neurosci. 9, 971-978. doi: 10.1038/nn1727
PubMed Abstract | CrossRef Full Text | Google Scholar
Wong, S. W., Massé, N., Kimmerly, D. S., Menon, R. S., e Shoemaker, J. K. (2007). Ventral medial prefrontal córtex e controle cardiovagal em humanos conscientes. Neuroimage 35, 698-708. doi: 10.1016/j.neuroimage.2006.12.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Yanaka, H. T., Saito, D. N., Uchiyama, Y., e Sadato, N. (2010). Substratos neurais de alerta de fase: um estudo de ressonância magnética funcional. Neurosci. Res. 68, 51-58. doi: 10.1016/j.neures.2010.05.005
PubMed Abstract | CrossRef Full Text | Google Scholar